Aminoácido non proteinoxénico
Os aminoácidos non proteinoxénicos ou non codificados (ou, ás veces, non estándar) son aminoácidos que non están codificados no código xenético. Ademais dos 23 aminoácidos codificados no código xenético (2 deles codificados dun modo especial e a N-formilmetionina que é un derivado de aminoácido), que se incorporan ás proteínas durante a tradución de proteínas nos ribosomas, e que se chaman aminoácidos proteinoxénicos, existen uns 140 aminoácidos naturais máis e coñécense miles de combinacións posibles máis, algúns dos cales poden aparecer formando parte das proteínas e outros non.[1] Algúns aminoácidos non proteinoxénicos son importantes porque:
- son intermediatos nas biosínteses
- incorpóranse postraducionalmente ás proteínas
- teñen unha función fisiolóxica (por exemplo, os compoñentes dos peptidoglicanos das paredes celulares bacterianas, neurotransmisores e toxinas)
- son compostos farmacolóxicos naturais ou producidos polo ser humano
- poden ser incorporados aos péptidos non ribosómicos, que non se sintetizan nos ribosomas
- detéctase a súa presenza en meteoritos e en experimentos prebióticos como o experimento de Miller e Urey.
Tipos de aminoácidos
editar
Todos os compostos orgánicos que levan un grupo funcional amino (-NH2) e un grupo carboxilo (-COOH) son aminoácidos. Os aminoácidos proteinoxénicos son un pequeno grupo de aminoácidos que teñen un átomo de carbono, que é o carbono 2 ou α, ao que están unidos o grupo amino, o grupo carboxilo, unha cadea lateral variable e un átomo de hidróxeno α, e que teñen unha conformación levo (L), coa excepción da glicina, que é aquiral (a prolina é considerada pola IUPAC un aminoácido, aínda que ás veces nalgúns libros aínda o consideren iminoácido).
O código xenético codifica 20 aminoácidos estándar. Porén, hai outros tres aminoácidos proteinoxénicos adicionais: selenocisteína, e pirrolisina e N-formilmetionina (un derivado de aminoácido polo que nas bacterias se inicia a síntese proteica). Os dous primeiros non teñen un codón específico, pero engádense en lugar do codón de finalización cando está presente unha secuencia específica, o codón UGA e o elemento SECIS para a selenocisteína, e o codón UAG e a secuencia PYLIS para a pirrolisina.[2][3] A formilmetionina está codificada nas bacterias polo codón AUG, que é á vez o codón de iniciación e o codón da metionina nas arqueas e eucariotas (pero en mitocondrias e cloroplastos é a N-formilmetionina). Un ARNt especial leva a N-formilmetionina ao codón AUG. Normalmente, a N-formilmetionina é eliminada posteriormente por modificación postraducional, pero non sempre. Tecnicamente a N-formilmetionina é un derivado de aminoácido (xa que o formilo está unido precisamente ao grupo amino) máis que un aminoácido normal e as máis das veces é eliminada despois da tradución, polo que ás veces non se conta como aminoácido proteinoxénico, pero a súa codificación é, basicamente, coma a doutro aminoácido calquera.
-
 Formilmetionina. Este derivado de aminoácido é unha metionina cuxo grupo amino foi protexido pola unión dun grupo formilo.
Formilmetionina. Este derivado de aminoácido é unha metionina cuxo grupo amino foi protexido pola unión dun grupo formilo. -
 Selenocisteína. Este aminoácido contén un grupo selenol no seu carbono β.
Selenocisteína. Este aminoácido contén un grupo selenol no seu carbono β. -
 Pirrolisina. Este aminoácido está formado pola unión do grupo ε-amino da lisina cun anel de pirrolina carboxilado.
Pirrolisina. Este aminoácido está formado pola unión do grupo ε-amino da lisina cun anel de pirrolina carboxilado.
-N-Formylmethionine_V.1.svg)

Os aminoácidos poden clasificarse en varios grupos:[4]
- Os 20 aminoácidos estándar normais. Codificados de forma normal no código xenético.
- Os 23 aminoácidos proteinoxénicos (22 se non contamos a N-formilmetionina). Son os 20 estándar normais máis 3 que están codificados dun modo especial.
- uns 80 aminoácidos creados abioticamente en altas concentracións.
- uns 900 que son producidos en vías biosintéticas naturais.
- uns 118 aminoácidos engadidos artificialmente ás proteínas.
Estes grupos solápanse pero non son idénticos. Os aminoácidos proteinoxénicos son biosintetizados polos organismos vivos, pero non todos son abióticos (non se encontran ademais en experimentos prebióticos e meteoritos), como a histidina. Moitos aminoácidos, como a ornitina, son intermediatos metabólicos que son bióticos, pero non están codificados. Outros son só intermediatos metabólicos, como a citrulina. Outros só aparecen en mesturas abióticas, como a α-metilnorvalina. Artificialmente, foron inseridos traducionalmente en proteínas uns 30 aminoácidos non naturais en sistemas de enxeñaría de proteínas, que non son biosintéticos.[4]
Nomenclatura
editarAdemais do sistema de numeración da nomenclatura da IUPAC para diferenciar os carbonos das moléculas orgánicas asignándolles un número, os carbonos tamén se nomean con letras gregas. No sistema de letras gregas o carbono α é o carbono central quiral que leva o grupo carboxilo (ao carbono do grupo carboxilo non se lle asigna letra grega), e os seguintes carbonos noméanse con letras gregas pola orde do alfabeto grego. Un α-aminoácido, ten o seu grupo amino unido ao carbono α (carbono 2), un β-aminoácido teno unido ao carbono β (C3) etc.[5]
Naturais, pero non L-α-aminoácidos
editarA maioría dos aminoácidos naturais son α-aminoácidos coa conformación L, pero existen algunhas excepcións.
Non alfa
editar
Existen algúns aminoácidos non α importantes, como a β-alanina (un β-aminoácido) e o GABA (un γ-aminoácido).
A β-alanina prodúcea a aspartato descarboxilase e é unida ao pantoato por medio dun enlace amida, formando ácido pantoténico, un precursor do coencima A.[6]
O GABA (ácido γ-aminobutírico) é un neurotransmisor co grupo amino no extremo da cadea lateral e un carbono máis comparado coa β-alanina.
Outros son: o ácido δ-aminolevulínico, que é un intermediato na biosíntese de tetrapirrois (hemo, clorofila, cobalamina etc.), e o ácido 4-aminobenzoico (PABA), que é un intermediato na biosíntese do folato.
-
 β-alanina
β-alanina -
 Ácido γ-aminobutírico (GABA)
Ácido γ-aminobutírico (GABA) -

-
 Ácido 4-aminobenzoico (PABA)
Ácido 4-aminobenzoico (PABA)



A razón pola que son os α-aminoácidos os que se utilizan nas proteínas atribúese á súa grande frecuencia nos meteoritos e experimentos prebióticos, polo que se supón que eran abundantes no momento da orixe da vida.[7] Unha especulación inicial que se formulou sobre as propiedades deletéreas dos β-aminoácidos na estrutura secundaria das proteínas,[7] comprobouse que era incorrecta.[8] Ademais existen varios inhibidores producidos artificialmente que non son α-aminoácidos, como a isoserina.
D-aminoácidos
editarA maioría das paredes das células bacterianas están formadas por peptidoglicano, un polímero composto por cadeas dun polisacárido enlazadas transversalmente con curtas pontes de oligopéptidos. Os oligopéptidos non se sintetizan nos ribosomas e conteñen varias peculiaridades, como a presenza de D-aminoácidos, xeralmente D-alanina e D-glutamato. Outra singularidade é que o primeiro é recemizado por encimas de unión ao piridoxal fosfato ou PLP (codificadas polos xenes alr ou o homólogo dadX), mentres que o segundo é racemizado por un encima independente de cofactores (murI). Preséntanse algunhas variantes en bacterias Thermotoga spp, nas cales está presente a A D-lisina, e en certas cepas resistentes á vancomicina aparece a D-serina.[9][10]
En animais, algúns D-aminoácidos son neurotransmisores.
Sen hidróxeno no carbono α
editarTodos os aminoácidos proteinoxénicos teñen polo menos un hidróxeno no seu carbono α: isto débese á especificidade da actividade transferase ribosómica e aos problemas biosintéticos do carbono cuaternario, que bloquearían a catálise dependente de PLP.[7]
Porén, hai algunhas excepcións. Nalgúns fungos o ácido α-amino isobutírico prodúcese como monómero para a síntese de certos antibióticos.[11] Este composto é similar á alanina, pero posúe un grupo metilo en lugar do hidróxeno do carbono α, polo que posúe dous grupos metilo no carbono α (un deles o da cadea lateral), polo que o carbono α non é un estereocentro. Outro composto similar á alanina é a deshidroalanina, que presenta unha cadea lateral meteno.
-
 Alanina
Alanina -
 Ácido aminoisobutírico
Ácido aminoisobutírico -
 Deshidroalanina
Deshidroalanina



Estereocentros de aminoácidos xemelgos
editarUn grupo de L-α-aminoácidos posúe dous extremos que poden considerarse α-aminoácidos (aínda que só un deles se considerará extremo α). Nas proteínas os residuos de cisteína forman entre eles unha unión por ponte disulfuro formando enlaces entre partes dunha proteína ou de distintas cadeas proteicas. Estes dímeros de cisteína denomínanse cistinas. A cisteína e a metionina prodúcense normalmente por sulfurilación directa, pero en certas especies poden producirse por transfurilación, na cal unha homoserina ou serina activadas se fusiona cunha cisteína ou homocisteína formando cistationina, unha molécula composta por un resto serina/cisteína unido por unha ponte tioéter cunha homoserina/homocisteína. Un composto natural similar é a lantionina, a cal pode considerarse como dúas alaninas unidas por un tióeter (ou dúas cisteínas monosulfuro, en vez de disulfuro como na cistina). De xeito similar, o ácido djenkólico, unha toxina da planta fabácea jengkol (Archidendron pauciflorum), está composta por dúas cisteínas unidas por dous tioéteres separados por un grupo metileno. O ácido diaminopimélico utilízase como ponte nos peptidoglicanos ou como precursor da lisina (por medio da súa descarboxilación).
-
 Cistina
Cistina -
 Cistationina
Cistationina -
 Lantionina
Lantionina -
 Ácido djenkólico
Ácido djenkólico -
 Ácido diaminopimélico
Ácido diaminopimélico

Funcións
editarNas células, especialmente as autótrofas, varios aminoácidos non proteinoxénicos son intermediatos metabólicos. Porén, a pesar da flexibilidade catalítica dos encimas que se unen ao PLP, moitos aminoácidos sintetízanse como cetoácidos (por exemplo, de 4-metil-2-oxopentanoato a leucina) e son aminados no último paso, o que mantén a cantidade de aminoácidos non proteinoxénicos intermediatos bastante baixa.
A ornitina e citrulina interveñen no ciclo da urea, que é parte do catabolismo de aminoácidos.[12]
Ademais de interviren no metaboismo primario, varios aminoácidos non proteinoxénicos son precursores para a produción final no metabolismo secundario de compostos como certas toxinas.
Aminoácidos prebióticos e do espazo
editarEn meteoritos caídos na Terra e en experimentos prebióticos (por exemplo, o experimento de Miller e Urey) atopáronse moitos máis aminoácidos ca os 20 aminoácidos estándar, varios dos cales en maiores concentracións ca os estándar, o que permitiu conxecturar que se a vida baseada nos aminoácidos se orixinou paralelamente tamén noutras partes do Universo, esta non tería en común coa vida terrestre máis do 75% dos aminoácidos.[7] A anomalía máis notable é a falta de ácido aminobutírico.
| Proporción de aminoácidos en relación á glicina (%) | ||
|---|---|---|
| Molécula | Descarga eléctrica | meteorito de Murchison |
| Glicina | 100 | 100 |
| Alanina | 180 | 36 |
| Ácido α-amino-n-butírico | 61 | 19 |
| Norvalina | 14 | 14 |
| Valina | 4.4 | |
| Norleucina | 1.4 | |
| Leucina | 2.6 | |
| Isoleucina | 1.1 | |
| Aloisoleucina | 1.2 | |
| t-Leucina | < 0.005 | |
| Ácido α-amino-n-heptanoico | 0.3 | |
| Prolina | 0.3 | 22 |
| Ácido pipecólico | 0.01 | 11 |
| Ácido α,β-diaminopropiónico | 1.5 | |
| Ácido α,γ-diaminobutírico | 7.6 | |
| Ornitina | < 0.01 | |
| Lisina | < 0.01 | |
| Ácido aspártico | 7.7 | 13 |
| Ácido glutámico | 1.7 | 20 |
| Serina | 1.1 | |
| Treonina | 0.2 | |
| Alotreonina | 0.2 | |
| Metionina | 0.1 | |
| Homocisteína | 0.5 | |
| Homoserina | 0.5 | |
| β-Alanina | 4.3 | 10 |
| Ácido β-amino-n-butírico | 0.1 | 5 |
| Ácido β-aminoisobutírico | 0.5 | 7 |
| Ácido γ-aminobutírico | 0.5 | 7 |
| Ácido α-aminoisobutírico | 7 | 33 |
| Isovalina | 1 | 11 |
| Sarcosina | 12.5 | 7 |
| N-etil glicina | 6.8 | 6 |
| N-propil glicina | 0.5 | |
| N-isopropil glicina | 0.5 | |
| N-metil alanina | 3.4 | 3 |
| N-etil alanina | < 0.05 | |
| N-metil β-alanina | 1.0 | |
| N-etil β-alanina | < 0.05 | |
| Isoserina | 1.2 | |
| Ácido α-hidroxi-γ-aminobutírico | 17 |
Cadea lateral recta
editarO código xenético foi descrito como un accidente conxelado durante a evolución inicial da vida e as razóns polas que hai só un aminoácido estándar cunha cadea lateral recta (a alanina) podería ser simplemente unha redundancia coa valina, leucina e isoleucina.[7] Porén, os aminoácidos de cadea lateral recta poden formar hélices alfa moito máis estables, polo que non terían nigún impedimento para formar parte das proteínas.[13]
-
 Glicina (a cadea lateral é un hidróxeno)
Glicina (a cadea lateral é un hidróxeno) -
Alanina (a cadea lateral é un metilo)
-
 Ácido α-aminobutírico (a cadea lateral é un etilo)
Ácido α-aminobutírico (a cadea lateral é un etilo) -
 Norvalina (a cadea lateral é un n-propil)
Norvalina (a cadea lateral é un n-propil) -
 Norleucina (a cadea lateral é un n-butil)
Norleucina (a cadea lateral é un n-butil) -
 Homonorleucina (a cadea lateral é un n-pentil)
Homonorleucina (a cadea lateral é un n-pentil)




Calcóxenos
editarOs calcóxenos son os elementos da táboa periódica da columna do oxíxeno (como oxíxeno, xofre, selenio, e telurio), que poden formar parte tamén das cadeas laterais de aminoácidos non proteinoxénicos e proteinoxénicos.
A serina, homoserina, O-metil-homoserina e O-etil-homoserina posúen cadeas laterais que son, respectivamente, un hidroximetil, hidroxietil, O-metil-hidroximetil e O-metil-hidroxietil. Mentres que a cisteína, homocisteína, metionina e etilonina posúen os equivalentes tiol. Os equivalentes selenol son a selenocisteína, selenohomocisteína, selenometionina e selenoetionina. Os aminoácidos co seguinte calcóxeno tamén se atopan na natureza: varias especies de fungos como Aspergillus fumigatus, Aspergillus terreus, e Penicillium chrysogenum en ausencia de xofre poden producir e incorporar ás proteínas telurocisteína e telurometionina.[14]
A hidroxiglicina, un aminoácido cunha cadea lateral hidroxilo, é moi inestable.
Código xenético expandido
editarDesde o ano 2001, foron engadidos uns 40 aminoácidos non naturais a proteínas creando para iso un codón especial (recodificación) e o correspondente par ARNt-aminoacil-ARNt-sintetase para codificalo. Utilízanse como ferramentas para estudar a estrutura das proteínas e o seu funcionmento ou para crear proteínas novas ou melloradas. Un código xenético con este tipo de modificacións chámase código xenético expandido.[15][16]
Incorporados ás proteínas postraducionalmente
editar- Véxase tamén: modificación postraducional.
Malia non estar codificados no código xenético, algúns aminoácidos non proteinoxenéticos aparecen incorporados ás proteínas. Orixínanse por modificación postraducional das cadeas laterais de aminoácidos estándar presentes na proteína. Estas modificacións son a miúdo esenciais para o funcionamento e regulación da proteína. Por exemplo, no gamma-carboxiglutamato a carboxilación do glutamato permite que se produza unha unión máis doada de catións calcio,[17] e na hidroxiprolina a hidroxilación da prolina é fundamental para a estabilidade do coláxeno e o mantemento do tecido conectivo.[18] Outro exemplo é a formación de hipusina no factor de iniciación eucariótico da tradución EIF5A, por modificación dun residuo de lisina.[19] Tales modificacións poden tamén determinar a localización da proteína, por exemplo, a adición de longos grupos hidrofóbicos pode causar que unha proteína se una aos fosfolípidos das membranas.[20]
-
 Ácido carboxiglutámico. O ácido glutámico posúe un grupo γ-carboxilo, e o ácido carboxiglutámico posúe dous.
Ácido carboxiglutámico. O ácido glutámico posúe un grupo γ-carboxilo, e o ácido carboxiglutámico posúe dous. -
 Hidroxiprolina. Difire da prolina no seu grupo hidroxilo no carbono 4.
Hidroxiprolina. Difire da prolina no seu grupo hidroxilo no carbono 4. -
 Hipusina. Este aminoácido obtense por adición ao grupo ε-amino dunha lisina dun residuo 4-aminobutil (obtido da espermidina.)
Hipusina. Este aminoácido obtense por adición ao grupo ε-amino dunha lisina dun residuo 4-aminobutil (obtido da espermidina.)


Análogos tóxicos
editarVarios aminoácidos non proteinoxénicos son tóxicos debido á súa capacidade de imitar certas propiedades dos aminoácidos proteinoxénicos, como a tialisina. Algúns aminoácidos non proteinoxénicos son neurotóxicos ao imitaren a aminoácidos que funcionan como neurotransmisores (é dicir, non para a biosíntese de proteínas), por exemplo, o ácido quiscuálico, a canavanina ou o ácido azetidina-2-carboxílico.[21] A cefalosporina C ten un esqueleto de ácido α-aminoadípico (homoglutamato) que está amidado cun resto de cefalosporina.[22] A penicilamina é un aminoácido terapéutico, cuxo modo de acción non se coñece. As cianotoxinas son compostos naturais que poden tamén incluír aminoácidos non proteinoxénicos. A microcistina e a nodularina, por exemplo, derivan ambas do β-aminoácido ADDA.
-
 Tialisina.
Tialisina. -
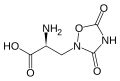 Ácido quiscuálico.
Ácido quiscuálico. -
 Canavanina.
Canavanina. -
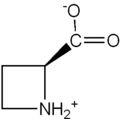 Áciddo azetidina-2-carboxílico.
Áciddo azetidina-2-carboxílico. -
 Cefalosporina C.
Cefalosporina C. -
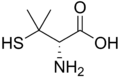 Penicilamina.
Penicilamina.

-Azetidine-2-carboxylate.png)

Os que non son aminoácidos
editarA taurina, que ás veces se require en cantidades relativamente grandes na dieta de certos animais, como os gatos (auxotrofia), non é un aminoácido senón un ácido aminosulfónico. Os osmólitos sarcosina e glicina betaína derivan de certos aminoácidos, pero teñen unha amina secundaria e cuaternaria, respectivamente.
Notas
editar- ↑ Ambrogelly, A.; Palioura, S.; Söll, D. (2007). "Natural expansion of the genetic code". Nature Chemical Biology 3 (1): 29–35. doi:10.1038/nchembio847. PMID 17173027.
- ↑ Böck, A.; Forchhammer, K.; Heider, J.; Baron, C. (1991). "Selenoprotein synthesis: An expansion of the genetic code". Trends in biochemical sciences 16 (12): 463–467. doi:10.1016/0968-0004(91)90180-4. PMID 1838215.
- ↑ Théobald-Dietrich, A.; Giegé, R.; Rudinger-Thirion, J. L. (2005). "Evidence for the existence in mRNAs of a hairpin element responsible for ribosome dependent pyrrolysine insertion into proteins". Biochimie 87 (9–10): 813–817. doi:10.1016/j.biochi.2005.03.006. PMID 16164991.
- ↑ 4,0 4,1 Lu, Y.; Freeland, S. (2006). "On the evolution of the standard amino-acid alphabet". Genome Biology 7 (1): 102. doi:10.1186/gb-2006-7-1-102. PMC 1431706. PMID 16515719.
- ↑ Voet, D. and Voet, J. G., Biochemistry (3rd ed.), John Wiley & Sons (2004)
- ↑ Coxon KM; Chakauya E; Ottenhof HH; et al. (2005). "Pantothenate biosynthesis in higher plants". Biochemical Society Transactions 33 (Pt 4): 743–6. PMID 16042590. doi:10.1042/BST0330743.
- ↑ 7,0 7,1 7,2 7,3 7,4 Weber, A. L.; Miller, S. L. (1981). "Reasons for the occurrence of the twenty coded protein amino acids". Journal of molecular evolution 17 (5): 273–284. doi:10.1007/BF01795749. PMID 7277510.
- ↑ Koyak MJ, Cheng RP. Design and synthesis of biologically active β-peptides. Meth Mol Biol.
- ↑ Boniface, A.; Parquet, C.; Arthur, M.; Mengin-Lecreulx, D.; Blanot, D. (2009). "The Elucidation of the Structure of Thermotoga maritima Peptidoglycan Reveals Two Novel Types of Cross-link". Journal of Biological Chemistry 284 (33): 21856–21862. doi:10.1074/jbc.M109.034363. PMC 2755910. PMID 19542229.
- ↑ Arias, C. A.; Martín-Martinez, M.; Blundell, T. L.; Arthur, M.; Courvalin, P.; Reynolds, P. E. (1999). "Characterization and modelling of VanT: A novel, membrane-bound, serine racemase from vancomycin-resistant Enterococcus gallinarum BM4174". Molecular microbiology 31 (6): 1653–1664. doi:10.1046/j.1365-2958.1999.01294.x. PMID 10209740.
- ↑ Gao, X.; Chooi, Y. H.; Ames, B. D.; Wang, P.; Walsh, C. T.; Tang, Y. (2011). "Fungal Indole Alkaloid Biosynthesis: Genetic and Biochemical Investigation of the Tryptoquialanine Pathway in Penicillium aethiopicum". Journal of the American Chemical Society 133 (8): 2729–2741. doi:10.1021/ja1101085. PMC 3045477. PMID 21299212.
- ↑ Curis E; Nicolis I; Moinard C; et al. (2005). "Almost all about citrulline in mammals". Amino Acids 29 (3): 177–205. PMID 16082501. doi:10.1007/s00726-005-0235-4.
- ↑ Padmanabhan, S.; Baldwin, R. L. (1991). "Straight-chain non-polar amino acids are good helix-formers in water". Journal of molecular biology 219 (2): 135–137. doi:10.1016/0022-2836(91)90553-I. PMID 2038048.
- ↑ Ramadan, S. E.; Razak, A. A.; Ragab, A. M.; El-Meleigy, M. (1989). "Incorporation of tellurium into amino acids and proteins in a tellurium-tolerant fungi". Biological trace element research 20 (3): 225–232. doi:10.1007/BF02917437. PMID 2484755.
- ↑ Xie J, Schultz PG (2005). "Adding amino acids to the genetic repertoire". Curr Opin Chem Biol 9 (6): 548–54. PMID 16260173. doi:10.1016/j.cbpa.2005.10.011.
- ↑ Wang Q, Parrish AR, Wang L (2009). "Expanding the genetic code for biological studies". Chem. Biol. 16 (3): 323–36. PMC 2696486. PMID 19318213. doi:10.1016/j.chembiol.2009.03.001.
- ↑ Vermeer C (1990). "Gamma-carboxyglutamate-containing proteins and the vitamin K-dependent carboxylase". The Biochemical Journal 266 (3): 625–36. PMC 1131186. PMID 2183788.
- ↑ Bhattacharjee A, Bansal M (March 2005) (2005). "Collagen structure: the Madras triple helix and the current scenario". IUBMB Life 57 (3): 161–72. PMID 16036578. doi:10.1080/15216540500090710.
- ↑ Park MH (2006). "The post-translational synthesis of a polyamine-derived amino acid, hypusine, in the eukaryotic translation initiation factor 5A (eIF5A)". Journal of Biochemistry 139 (2): 161–9. PMC 2494880. PMID 16452303. doi:10.1093/jb/mvj034.
- ↑ Blenis J, Resh MD (1993). "Subcellular localization specified by protein acylation and phosphorylation". Current Opinion in Cell Biology 5 (6): 984–9. PMID 8129952. doi:10.1016/0955-0674(93)90081-Z.
- ↑ Dasuri, K.; Ebenezer, P. J.; Uranga, R. M.; Gavilán, E.; Zhang, L.; Fernandez-Kim, S. O. K.; Bruce-Keller, A. J.; Keller, J. N. (2011). "Amino acid analog toxicity in primary rat neuronal and astrocyte cultures: Implications for protein misfolding and TDP-43 regulation". Journal of Neuroscience Research 89 (9): 1471–1477. doi:10.1002/jnr.22677. PMC 3175609. PMID 21608013.
- ↑ Trown, P. W.; Smith, B.; Abraham, E. P. (1963). "Biosynthesis of cephalosporin C from amino acids". The Biochemical journal 86 (2): 284–291. PMC 1201751. PMID 13994319.