エストロゲン受容体
エストロゲン受容体(エストロゲンじゅようたい、英:Estrogen Receptor、ER)とはステロイド受容体スーパーファミリーに属する分子の一つである。卵胞ホルモン受容体とも呼ばれる。そもそもエストロゲンとはエストロン(E1)、エストラジオール(E2)およびエストリオール(E3)の3種類の分子を指しており、いずれもERとの結合能を有するが、中でも生体における産生量はE2が多い。エストロゲンはステロイドホルモンの一種であり、生殖機能の形成および細胞の増殖を促進する働きを持つ。その生理作用を発現するためには標的組織に存在しているERへの結合を介する必要がある。ERに対してリガンドが結合するとERは活性化[1]を受けてDNAへの結合が促進され、遺伝子の転写を制御する転写因子として機能する。また、植物中に含まれるイソフラボンなどの分子(植物性エストロゲン)や内分泌撹乱物質もERに対して結合能を有し、作用を発現することが知られている。
研究の経緯
編集
1950年代にエルウッド・ジェンセンとハーバート・ヤコブソンは放射性元素であるトリチウムでラベルしたエストロゲン分子が標的組織に集積する性質を持つことを見出した。そこで、これらの組織にはエストロゲンに対する受容体が存在するのではないかという仮説に行き当たり、1966年にヒトやラットの組織からエストロゲン受容体(ER)が発見された(後にERαと呼ばれる分子である)。その後、1986年にERをコードする遺伝子配列が分かり、1993年にER遺伝子のノックアウトマウスが作成された。1996年に第二のERタンパク質であるERβがラットの前立腺から発見され、その解釈は複雑さを増した。研究が進むと、ERαとERβの間にはDNA結合ドメインの相同性もなく、組織分布も異なることが分かり、それぞれ異なった機能を有していることが示されている。
構造
編集核内受容体
編集
ERには2つのアイソフォームが存在しており、それぞれERα(NR3A1、595アミノ酸残基)およびERβ(NR3A2、530残基)と呼ばれる。これらは独立した遺伝子(ESR1、ESR2)から産生され、選択的スプライシングによる産物ではない。ESR1は6q25.1に存在し、ESR2は14q21-22に存在している。リガンドの結合により活性化されたERタンパク質はホモ(αα、ββ)あるいはヘテロ二量体(αβ)を形成する。また、ERαとERβはいずれも6つのドメイン(A-F領域)から構成されている。2種類の受容体タンパク質間ではアミノ酸配列の配列類似性が高く、DNA結合領域(C領域)で96%、リガンド結合領域(E/F領域)は若干低く58%となっているが、A/B領域やD領域では配列類似性が低い。ERの転写活性化に関与する2つのドメイン構造はA/B領域およびE領域内に存在し、それぞれAF-1およびAF-2と呼ばれている。これらのリガンドに対する反応性はそれぞれ異なり、AF-1による転写活性化はリガンド非依存性であり恒常的に転写活性化能を示すが、AF-2による転写活性化はリガンドの結合に依存している。さらに、DNA結合領域(C領域)には2つのZnフィンガーモチーフが含まれ、DNA上に存在する応答エレメント(ERE:AGGTCAnnnTGACCT、nはATGCいずれかの核酸)との結合に関与している。D領域はヒンジ領域と呼ばれ、受容体タンパク質の柔軟性を形成している。カルボキシル基末端側のEおよびF領域はリガンドとの結合に関与しているドメインであり、リガンド非結合状態においてはhsp90やhsp70等の分子シャペロンと結合している。また、この領域は受容体タンパク質の二量体形成においても重要な働きをしている。
Gタンパク質共役受容体
編集
1975年、PietrasとSzegoによって細胞膜上にエストロゲンの結合部位が存在し、早いシグナル伝達を引き起こすことが報告された[2][3]。この早いシグナル伝達は生殖関連組織である子宮や卵巣をはじめ、乳房、骨、神経組織において見られる。近年、オーファン受容体であるGPR30が膜上に存在するエストロゲン受容体であり、このシグナル伝達に関与している可能性が示されている[4]。GPR30はGタンパク質共役受容体の一種であり、核内受容体タイプのERがリガンドの結合により転写因子としてDNAと相互作用を行うのに対して、GPR30はDNAとの直接的な相互作用を行わず、下流に存在するセカンドメッセンジャーを介してイオンチャネルに対して影響を与えるなどの遺伝子に非依存的な経路も存在する。また、乳癌の細胞においてはGPR30からの刺激が核内ERのリン酸化を引き起こし、転写活性化を引き起こすことも報告されている[5]。しかし、2008年現在においてGPR30が関与する生理機能については未だ明らかではない。
リガンドおよび阻害薬の例
編集リガンド
編集阻害薬
編集- タモキシフェン(Tamoxifen)
- ラロキシフェン(Raloxifen)
- ICI182,780
-
 エストロン
エストロン -
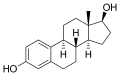 17β-エストラジオール
17β-エストラジオール -
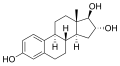 エストリオール
エストリオール -
 タモキシフェン
タモキシフェン




転写活性化機構
編集従来ERの活性化のスイッチはリガンドの結合であると考えられてきた。リガンドの結合はERの二量体の形成とそれに引き続いて起こる核内移行を引き起こす。ERはさらにDNA上のエンハンサー配列であるEREに対して結合して遺伝子の転写活性化させ、その作用を発現するというのが古典的なモデルであった。確かにリガンドの結合によって転写活性化が引き起こされることには間違いないのだが、近年の研究ではERを含むステロイド受容体の活性化はリガンドの結合よりもむしろ翻訳後修飾が重要な役割を担っていることが分かってきており、リガンドの結合を必要としない転写活性化経路も存在する。この翻訳後修飾を「受容体の活性化」と呼ぶことがあり、例えばERのリン酸化はリガンド依存的・非依存的のいずれの転写活性化機構も促進し、中でもアミノ基末端側から118番目のセリン残基(S118)のMAPキナーゼ[6]やCDK(サイクリン依存性キナーゼ)7[7]によるリン酸化が重要であることが報告されている。また、転写活性化には単にERがDNAに結合すればよいというわけではなく、そのほかにも多くの分子が関与する複雑な機構であることが分かってきている。これらのタンパク質との結合部位となるのが既述のAF-1およびAF-2ドメインであり、AF-1はリガンド非依存的な転写活性化機構に関与しており、その転写活性の強さは受容体のリン酸化状態と密接な関係がある。一方、AF-2を介した転写活性化はリガンド依存的であることが知られる。AF-2の機能にはリガンド結合ドメインのカルボキシル基末端側に位置するHelix12とよばれる短いらせん状の構造が重要である。
抑制的制御機構
編集リガンドであるエストロゲンがERに結合すると遺伝子の転写活性化が引き起こされる一方で、ERを介した反応を抑制的に制御するフィードバックが働き、下記に示すようないくつかの機構が知られている。
機能および疾患への関与
編集エストロゲンは子宮や卵巣、乳腺、前立腺、肺、脳などの形態形成に関与していることが報告されている一方、ノックアウトマウスの解析も行われており、ERは生存および胎生期における生殖器の形成に必ずしも必要ではないことが分かっている[11]。
エストロゲンはがん細胞の増殖を促進する作用を有しており、乳癌や子宮癌の患者の組織においては健常人と比較してERの発現が上昇していることが報告されている[12]。そのため、乳癌の患者では抗エストロゲン療法が行われ、ER拮抗薬であるタモキシフェンやエストロゲンの生合成に関与する酵素であるアロマターゼの阻害薬などが用いられる。また、近年乳癌の治療薬として開発された抗体医薬であるトラスツズマブはHER2に対するヒト化モノクローナル抗体であるが、HER2はMAPキナーゼであるERK1/2を活性化し、ERのリン酸化を引き起こすことによりERのリガンド非依存的な転写を活性化させることが報告されている。
出典
編集- 今堀和友、山川民夫 編集 『生化学辞典 第4版』東京化学同人 2007年 ISBN 9784807906703
参考文献
編集- ^ Dahlman-Wright K, Cavailles V, Fuqua SA, Jordan VC, Katzenellenbogen JA, Korach KS, Maggi A, Muramatsu M, Parker MG and Gustafsson JA.(2006)"International Union of Pharmacology. LXIV. Estrogen receptors."Pharmacol.Rev. 58,773-81. PMID 17132854.
- ^ Pietras RJ and Szego CM.(1975)"Endometrial cell calcium and oestrogen action."Nature. 253,357-9. PMID 1167402
- ^ Pietras RJ and Szego CM.(1977)"Specific binding sites for oestrogen at the outer surfaces of isolated endometrial cells."Nature. 265,69-72. PMID 834244
- ^ Revankar CM, Cimino DF, Sklar LA, Arterburn JB and Prossnitz ER.(2005)"A transmembrane intracellular estrogen receptor mediates rapid cell signaling."Science. 307,1625-30. PMID 15705806
- ^ Filardo EJ, Quinn JA, Frackelton AR Jr and Bland KI.(2002)"Estrogen action via the G protein-coupled receptor, GPR30: stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptorto-MAPK signaling axis."Mol.Endocrinol. 16,70-84. PMID 11773440
- ^ Kato S, Endoh H, Masuhiro Y, Kitamoto T, Uchiyama S, Sasaki H, Masushige S, Gotoh Y, Nishida E and Kawashima H.(1995)"Activation of the estrogen receptor through phosphorylation by mitogen-activated protein kinase."Science. 270,1491-4. PMID 7491495
- ^ Chen D, Riedl T, Washbrook E, Pace PE, Coombes RC, Egly JM and Ali S.(2000)"Activation of estrogen receptor alpha by S118 phosphorylation involves a ligand-dependent interaction with TFIIH and participation of CDK7."Mol.Cell. 6,127-137. PMID 10949034
- ^ Tschugguel W, Dietrich W, Zhegu Z, Stonek F, Kolbus A and Huber JC.(2003)"Differential regulation of proteasome-dependent estrogen receptor alpha and beta turnover in cultured human uterine artery endothelial cells. J Clin Endocrinol Metab."J.Clin.Endocrinol.Metab. 88,2281-7. PMID 12727987
- ^ Kaneko KJ, Furlow JD and Gorski J.(1993)"Involvement of the coding sequence for the estrogen receptor gene in autologous ligand-dependent downregulation."Mol.Endocrinol. 7,879-88. PMID 8413312
- ^ Pinzone JJ, Stevenson H, Strobl JS and Berg PE.(2004)"Molecular and cellular determinants of estrogen receptor alpha expression."Mol.Cell.Biol. 24,4605-12. PMID 15143157
- ^ Lubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS and Smithies O.(1993)"Alteration of reproductive function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene."Proc.Natl.Acad.Sci.USA. 90,11162-6. PMID 8248223
- ^ [No authors listed](1998)"Tamoxifen for early breast cancer: an overview of the randomised trials. Early Breast Cancer Trialists' Collaborative Group."Lancet. 351,1451-67. PMID 9605801