
Plàncton
| No s'ha de confondre amb Plankton. |


Segons Victor Hensen (1887), el plàncton (del grec antic: πλαγκτός / plangtós, que significa 'errant', 'vagabund') és el conjunt d'organismes que viuen en aigua dolça, salobre i salada, generalment suspesos i aparentment passius: gàmetes, larves, animals no aptes per lluitar contra el corrent (petits crustacis planctònics, meduses i sifonòfors), plantes i algues microscòpiques.
El plàncton és la base de moltes xarxes tròfiques aquàtiques. És el principal aliment de les balenes.[1]
Alguns organismes són capaços de tenir un moviment independent i poden nedar centenars de metres verticalment en un sol dia (un comportament anomenat migració vertical diària). La seva posició horitzontal es determina principalment pels corrents circumdants.
Els organismes planctònics es defineixen a partir del seu nínxol ecològic i no segons criteris filogenètics i taxonòmics.
El fitoplàncton constitueix al voltant del 50% de la matèria orgànica produïda sobre el planeta Terra, però sembla que pateix una disminució constant des de finals del segle xx.
El zooplàncton contribueix amb els seus moviments verticals (cicles relacionats amb la llum i les estacions) a la barreja de capes d'aigua. Aquest és un aspecte de la bioturbació que es pot haver subestimat.[2][3] Aquests fenòmens també existeixen en l'aigua dolça (amb el moviment de les poblacions de dàfnies, per exemple).
Terminologia
[modifica]El nom plàncton (creat pel zoòleg alemany Victor Hensen)[4] deriva de l'adjectiu grec πλαγκτός (plagktós), que significa errant o vagabund.[5]
Actualment, la definició d'Hensen és vista com a incompleta perquè no inclou algunes coses; el plàncton designa a un conjunt d'organismes diferents i s'ha d'especificar de quin organisme (tàxon) es parla.
Els científics han proposat diferents termes per alguns dels organismes que viuen en el medi ambient aquàtic:
- el nècton és capaç de moure's de manera activa horitzontalment i/o verticalment, a vegades contra corrent, i controlar la seva posició: peixos, calamars, mamífers marins, etc. El conjunt format per plàncton i nècton s'anomena pélagos.
- el trípton, anomenat pseudoplàncton per Davis (1955),[6] que reuneix els elements suposats morts (necromassa) o d'origen mineral o orgànic (excrements, partícules de plàncton mort ...).
- el sèston, inclou totes les partícules de tota classe, vives o mortes, orgàniques o inorgàniques suspeses en aigua (sèston = trípton + plàncton). El sèston és format en particular per les excrecions del plàncton i d'altres organismes, inclosos els seus excrements.
- l'holoplàncton són organismes que passen tot el seu cicle biològic formant part del plàncton (per exemple, la majoria de les algues, copèpodes, salpes, i algunes meduses).
- el meroplàncton són organismes que només són planctònics durant part de la seva vida (en general l'etapa larval), i després es canvien ja sigui a una existència nectònica o bentònica (al fons de la mar). Exemples de meroplàncton inclouen les larves d'eriçons de mar, estrelles de mar, crustacis, cucs marins, i la majoria de peixos.[7]
- l'ictioplàncton és el conjunt d'ous i larves de nècton que presenten un comportament planctònic, el que significa que no poden nedar amb eficàcia pels seus propis mitjans sinó que han de deixar-se portar pels corrents oceànics.
La branca de la biologia que s'ocupa de l'estudi d'aquests organismes, incloent-hi les interrelacions i interdependències físiques, químiques i amb l'entorn biològic, es diu planctologia.[8]
Tipus de plàncton
[modifica].jpg)


El plàncton es divideix principalment en amplis grups funcionals o nivells tròfics:
- El fitoplàncton (del grec: φυτόν / phutón, que significa «planta»), o plàncton vegetal, és el punt de partida per a tota activitat biològica del mar, la base de totes les cadenes alimentàries aquàtiques. Utilitza l'energia solar per a produir matèria orgànica. A excepció del nanoplàncton i del picoplàncton, està present principalment en les capes superficials del mar (de 0 a 15 metres de profunditat). Determina la seva posició, en la superfície o entre dues aigües, depenent de la quantitat de llum que pot rebre, essencial per a la seva fotosíntesi. Es compon d'algues microscòpiques formades a partir d'una sola cèl·lula o de cèl·lules agrupades en cadenes. Es reprodueix per divisió cel·lular amb l'ajuda de la llum, del CO₂ i dels nutrients (minerals i elements variats i complexos, especialment fòsfor i nitrogen que provenen de la descomposició dels residus orgànics per bacteris).[9] Produeix grans quantitats d'oxigen, necessari per a la vida a l'aigua, i per l'intercanvi de gasos contribueix a l'oxigenació del planeta. Dins el fitoplàncton, els dos grups més grans, més abundants en termes d'espècies, són les diatomees i els dinoflagel·lats.
- El zooplàncton (del grec: ζῷον / Zoio, que significa «animal»), o plàncton animal, es compon de dos grups: holoplàncton i meroplàncton. D'acord amb el cicle de vida dels organismes, l'holoplàncton (o plàncton permanent) es reprodueix per aparellament. El meroplàncton (o plàncton temporal) està format per moltes espècies marines (com la llagosta, les ostres, els musclos…) que en algun moment de la seva existència passen per etapes larvàries molt complexes. Els crustacis copèpodes constitueixen més del 80% del zooplàncton. Durant la nit puja a la superfície per alimentar-se de fitoplàncton i durant el dia baixa a aigües més profundes. Així, el zooplàncton s'escapa dels depredadors i estalvia energia, ja que la temperatura és més baixa. Aquest moviment del zooplàncton, que contribueix a la barreja d'aigua i que varia la temperatura i la quantitat d'oxigen entre les capes d'aigua, es diu migració vertical. Alguns depredadors del plàncton segueixen aquests moviments. En alguns litorals i costes urbanitzades, és possible que algunes espècies de zooplàncton es puguin veure afectades per la contaminació lumínica.
- el bacterioplàncton està format per bacteris que tenen almenys una fase de via lliure en l'aigua.
- el micoplàncton, fongs i organismes similars als fongs, també són importants en el cicle de nutrients, com el bacterioplàncton.[10]
- El nanoplàncton (de 20 a 2 µm), picoplàncton (de 2 a 0,2 µm), i femtoplàncton o virioplàncton (principalment els virus marins) han sigut descoberts recentment i encara són una part poc coneguda de la biodiversitat marina. Aquestes categories inclouen moltes espècies que poden viure a gran profunditat, on la intensitat de llum és extremadament baixa. Algunes d'aquestes espècies semblen tenir lenta taxa de reproducció i una excepcionalment llarga vida útil (característica que es troben també en organismes més complexos de les profunditats, com alguns peixos d'aigües molt profundes).
Aquest esquema divideix la comunitat del plàncton en amplis grups de productors, consumidors i descomponedors. No obstant això, la determinació del nivell tròfic d'alguns plàncton no és senzill. Per exemple, encara que la majoria de dinoflagel·lats són o bé productors fotosintètics o consumidors heteròtrofs, moltes espècies són mixotrófiques depenent de les circumstàncies.
Les mides del plàncton
[modifica]El plàncton també es descriu sovint en termes de mida.[11] En general, s'utilitzen les següents divisions:
Grup Mida (ESD) Exemples Megaplàncton > 20 cm metazoa: meduses, ctenòfors, sàlpids, pirosomes, urocordats, cefalòpodes; amfípodes Macroplàncton 20 - 2 cm metazoa: pteròpods, quetògnats, eufausiacis (krill), meduses, ctenòfors, sàlpids, pirosomes, cefalòpodes, janthinidae (una família de gastròpodes), amfípodes Mesoplàncton 20 - 0,2 mm metazoa: copèpodes, meduses, cladòcers, ostràcodes, quetògnats, pteròpods, urocordats, heteròpodes Microplàncton 200 - 20 µm grans eucariotes protists: la major part del fitoplàncton, protozous, foraminífers, tintínnids, altres ciliats, rotífers, metazoa juvenil - crustàcis (copèpod nauplii) Nanoplàncton 20 - 2 µm petits eucariotes protists: petites diatomees, petits flagel·lats, dinoflagel·lats, algues daurades, algues verdes, algues verdes-grogues Picoplàncton 2 - 0,2 µm petits eucariotes protists: bacteris, chrysophyta Femtoplàncton < 0,2 µm virus marins
No obstant això, alguns d'aquests termes poden ser utilitzats amb diferents límits, especialment a l'extrem més petit. L'existència i la importància del nanoplàncton i el plàncton encara més petit es va descobrir durant la dècada de 1980, però es creu que constitueixen la major proporció de tot el plàncton en quantitat i en diversitat.
El microplàncton i els grups més petits són microorganismes i operen a baixos nombres de Reynolds, on la viscositat de l'aigua és molt més important que la seva massa o inèrcia.[12]
Distribució
[modifica]
El plàncton habita els oceans, mars, llacs i estanys. L'abundància local varia horitzontalment, verticalment i estacionalment. La principal causa d'aquesta variabilitat és la disponibilitat de llum. Tots els ecosistemes de plàncton són moguts per l'entrada de l'energia solar (però també per la quimiosíntesi), limitant la producció primària en les aigües superficials, i en les regions geogràfiques i estacions que tenen abundant llum.
Una variable secundària és la disponibilitat de nutrients. Tot i que les grans àrees dels oceans tropicals i subtropicals tenen abundant llum, experimenten un producció primària relativament baixa, ja que ofereixen nutrients limitats com ara nitrats, fosfats i silicats. Això és conseqüència del corrent oceànic a gran escala i l'estratificació de la columna d'aigua. En aquestes regions, la producció primària en general es produeix a major profunditat, encara que a un nivell reduït (a causa de la poca llum).
Tot i les concentracions de macronutrients importants, algunes regions oceàniques són improductives (les denominades regions HNLC).[13] El micronutrient ferro és deficient en aquestes regions i, a més, pot conduir a la formació de flors d'aigua de molts tipus de fitoplàncton.[14] L'oceà aconsegueix el ferro principalment a través de la deposició de pols a la superfície del mar. Paradoxalment, les àrees oceàniques adjacents a les terres improductives i àrides tenen abundant fitoplàncton (per exemple, l'est de l'oceà Atlàntic, on els vents alisis porten la pols des del desert del Sàhara, al nord d'Àfrica).
Mentre que el plàncton és més abundant en les aigües superficials, també pot viure al llarg de la columna d'aigua. A grans profunditats no es produeix la producció primària, i el zooplàncton i el bacterioplàncton consumeixen matèria orgànica de les aigües superficials que es va enfonsant. Aquest flux de material que s'enfonsa, anomenat neu marina, pot ser especialment alt després de l'acabament de les flors d'aigua.
S'ha detectat els rastres de plàncton en la superfície de l'aigua des de l'espai, des de l'Estació Espacial Internacional.[15]
Les adaptacions del plàncton a la vida pelàgica
[modifica]Tot i l'àmplia varietat de plàncton, algunes característiques generals donen una fisonomia particular als organismes planctònics, que estan relacionades amb el seu estil de vida en aigües obertes. De fet, aquest mode de vida requereix adaptacions que permeten als organismes evitar ser vists pels depredadors, ja que són incapaços de moure's voluntàriament a través de grans distàncies i per tant són presa fàcil, i mantenir-se en la columna d'aigua evitant enfonsar-se.
- Coloració: Els organismes planctònics, en general, no està pigmentatst i fins i tot tendeixen a la transparència. En els organismes pigmentats, la pigmentació es limita a òrgans específics i petits (per exemple, els plastidis o òrgans de la visió).
- Flotabilitat: Els organismes planctònics han de mantenir-se entre dues aigües i evitar enfonsar-se utilitzant el mínim d'energia possible. Per a això, han desenvolupat diverses estratègies que els permeten flotar, ja sigui reduint el seu pes o augmentar la seva superfície de flotació:
- Són rics en aigua amb un cos que pot estar format de substàncies gelatinoses. Alguns individus que pertanyen al megaplàncton poden tenir un contingut d'aigua més gran que 95%. En comparació, el cos humà conté 56%.
- Pocs tenen formacions esquelètiques. Els organismes que tenen un esquelet el tenen lleuger i resistent. Les diatomees, per exemple, tenen frústuls molt prims i quitina.
- Igual que tots els éssers vius, els organismes planctònics tenen una densitat més gran que la de l'aigua a causa de la constitució del citoplasma de la cèl·lula (que té una densitat entre 1,02 i 1,06), però alguns organismes tenen vacúols o flotadors plens de lípids o de gas amb una densitat menor que l'aigua.
- Altres fins i tot han canviat la composició iònica de les seves cèl·lules: tenen ions lleugers com ara clor (Cl) o potassi (K), que reemplacen el sulfat (SO₄) i magnesi (Mg) de més pes molecular.
- Tenen cossos que augmenten la seva superfície. Per exemple, algunes diatomees tenen filaments. Altres organismes es reuneixen en colònies (per exemple els sàlpids, que formen cadenes de fins a més de 10 m de llarg).
Aquestes adaptacions a vegades no són suficients per a evitar l'enfonsament, i alguns organismes les han complementat amb una activitat motora reduïda a través de cilis, flagels o contraccions del cos.
El cicle biològic
[modifica]
Segons la forma en que realitzen el cicle biològic, els organismes planctònics es poden dividir en dues grans categories: holoplàncton i meroplàncton.

Els organismes holoplanctònics passen tota la seva vida surant de forma passiva en una zona pelàgica i, per tant, en cada etapa de la seva vida contribueix a la densitat de la població planctònica. Són part de l'holoplàncton diferents tipus d'algues unicel·lulars i pluricel·lulars, protozous, els cnidaris (com els sifonòfors i alguns escifozous), gairebé tots els ctenòfors, els rotífers, diversos grups de crustacis, mol·luscs gastròpodes, alguns dels quetògnats, molts urocordats i l'holotúria Pelagothuria natatrix com a representant dels equinoderms.
Els organismes meroplanctònics viuen només una part de la seva vida en la fase planctònica, i posteriorment es canvien ja sigui a una existència nectònica o bentònica. El meroplàncton es compon principalment de gàmetes, larves d'animals i espores de plantes bentòniques i, per aquesta característica, la seva densitat i biodiversitat estan fortament influenciades per les diferents èpoques de reproducció de la població aquàtica. Exemples de meroplàncton són: les larves d'esponges i mol·luscs (que en la seva fase adulta es dipositen en la part inferior, convertint-se en bentònics), actínules, efires i l'etapa de medusa dels cnidaris, les diferents formes larvàries de peixos que en la seva forma adulta que es mouen en el nècton o el bentos. Els briozous, anèl·lids, artròpodes, equinoderms, urocordats i molts fílums d'animals també són presents al plàncton en diverses formes larvàries.
La nutrició
[modifica]
Mitjançant l'anàlisi de les diferents formes en què els organismes adquireixen l'energia necessària per al metabolisme, és possible identificar el plàncton en els tres principals nivells tròfics de la cadena alimentària: productors, consumidors i descomponedors.
La producció està al càrrec del fitoplàncton, format per diverses espècies d'algues monocel·lulars i pluricel·lulars i que, com tots els organismes autòtrofs fotosintètics (plantes terrestres), són capaços de sintetitzar la matèria orgànica utilitzant les substàncies inorgàniques dissoltes en l'aigua i la radiació solar com a font d'energia. Per aquesta característica, el fitoplàncton es troba en la base de la cadena alimentària en la gran majoria dels ecosistemes aquàtics
El paper dels consumidors és del zooplàncton, incloent els protozous i metazous. Són organismes heteròtrofs que necessàriament s'han d'alimentar d'altres organismes per a obtenir l'energia emmagatzemada en la matèria orgànica de les seves preses.
Finalment, però no menys important, és el nivell tròfic que ocupa el bacterioplàncton, format pels bacteris que suren en la columna d'aigua. Obtenen l'energia a través de la descomposició de la necromassa i tornen a l'entorn aquàtic les substàncies inorgàniques que tornaran a ser utilitzades pel fitoplàncton.
Cal assenyalar que aquest esquema de subdivisió no presenta els límits del tot clars. De fet, algunes espècies de protists tenen la capacitat d'actuar tant com a productors fotosintètics o com a consumidors heterotròfics, depenent de les circumstàncies (mixotròfia).
La producció
[modifica]La producció primària, realitzada pel fitoplàncton (algues planctòniques), depèn de la disponibilitat de nutrients (nitrogen, fòsfor i silici d'acord amb les espècies de fitoplàncton), la temperatura i la llum a l'aigua. La producció secundària està relacionada amb la biomassa del zooplàncton (plàncton animal) i la seva eficàcia de creixement.
La producció, en relació amb la biomassa, és més gran en aigua freda, més densa i rica en nutrients. Sovint també és major en les zones costaneres sotmeses a l'enriquiment de nutrients pels rius.
Malgrat l'augment de la producció al nord, al voltant dels pols, i malgrat algunes floracions espectaculars locals, l'activitat del plàncton sembla que ha disminuït globalment entre 1999 i 2006.
El plàncton és la base de moltes xarxes tròfiques.
La seva importància en els cicles biogeoquímics
[modifica]A part de ser la base de moltes cadenes alimentàries que donen un important suport comercial a la indústria pesquera, els ecosistemes de plàncton tenen un paper en els cicles biogeoquímics de molts elements químics importants, incloent el cicle de carboni de l'oceà.[16]
Principalment pel pasturatge del fitoplàncton, el zooplàncton proporciona carboni a la xarxa tròfica, ja sigui per la respiració per a proporcionar energia metabòlica, o per la mort com a biomassa o detrit. La matèria orgànica tendeix a ser més densa que l'aigua de mar, i com a resultat s'enfonsa en els ecosistemes de mar obert lluny de les costes, transportant el carboni juntament amb ella. Aquest procés es coneix com la bomba biològica, i és una de les raons que els oceans constitueixen el major embornal de carboni en la Terra.
Seria possible augmentar la captació de l'oceà de diòxid de carboni generat a través de les activitats humanes mitjançant l'augment de la producció de plàncton a través de la fertilització, principalment amb ferro com a micronutrient. No obstant això, aquesta tècnica no pot ser pràctica a gran escala. L'esgotament de l'oxigen a l'oceà i la producció de metà resultant (causada per l'excés de producció remineralitzant en la profunditat) és un desavantatge potencial.[17][18]
La variabilitat de la biomassa
[modifica]El creixement de les poblacions de fitoplàncton depèn dels nivells de llum i de la disponibilitat de nutrients. El factor principal que limita el creixement varia d'una regió a una altra en els oceans del món. En una escala més àmplia, el creixement de fitoplàncton en l'oligotròfic tròpic i en els girs subtropicals generalment està limitat pel subministrament de nutrients, mentre que la llum sovint limita el creixement del fitoplàncton en els girs subàrtics.
La variabilitat ambiental en múltiples escales influeix en els nutrients i la llum disponible per al fitoplàncton, i atès que aquests organismes formen la base de la cadena alimentària marina, aquesta variabilitat en el creixement del fitoplàncton influeix en els nivells tròfics superiors. Per exemple, a escales interanuals el nivells de fitoplàncton es baixen temporalment durant els períodes de El Niño, que influeixen en les poblacions de zooplàncton, peixos, aus marines i mamífers marins.
Els efectes de l'escalfament global en la població de fitoplàncton és una àrea de recerca activa. S'espera que els canvis en l'estratificació vertical de la columna d'aigua, la velocitat de les reaccions biològiques dependents de la temperatura, i el subministrament atmosfèric de nutrients pot tenir un impacte important en el futur en la productivitat del fitoplàncton.[19] A més, els canvis en la mortalitat del fitoplàncton a causa de les taxes de pasturatge del zooplàncton pot ser significatiu.
Les larves de peixos acabats de descloure són també plàncton durant uns dies, el temps que necessiten abans de nedar contra corrent.
-

-
 Un copèpode (Calanoida sp.) prop de l'Antàrtida, de 12 mm de llarg, de forma ovoide, translúcid i amb dues antenes llargues
Un copèpode (Calanoida sp.) prop de l'Antàrtida, de 12 mm de llarg, de forma ovoide, translúcid i amb dues antenes llargues -
 Krill antàrtic (Euphausia superba), probablement la bimomassa més gran d'una sola espècie al planeta
Krill antàrtic (Euphausia superba), probablement la bimomassa més gran d'una sola espècie al planeta -
 Krill del nord (Meganyctiphanes norvegica), l'intestí és de color vermell i s'alimenta de zooplàncton
Krill del nord (Meganyctiphanes norvegica), l'intestí és de color vermell i s'alimenta de zooplàncton -
 Tomopteris és un gènere de poliquets marins planctònics
Tomopteris és un gènere de poliquets marins planctònics -
 Un ctenòfor, es pot veure la «cinta transportadora» de les larves que creixen i l'ovari
Un ctenòfor, es pot veure la «cinta transportadora» de les larves que creixen i l'ovari -
 L'escuma marina és produïda pel plàncton.
L'escuma marina és produïda pel plàncton. -
 Aleví de salmó que ha sortit del sac vitel·lí
Aleví de salmó que ha sortit del sac vitel·lí -
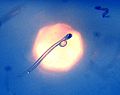 Imatge de la molt petita larva d'arengada amb les restes del sac vitel·lí i el llarg intestí visible en l'animal transparent
Imatge de la molt petita larva d'arengada amb les restes del sac vitel·lí i el llarg intestí visible en l'animal transparent -

-
 Larva de peix-gel en l'Antàrtida. No té hemoglobina
Larva de peix-gel en l'Antàrtida. No té hemoglobina -
 Microzooplancton, els principals depredadors del plàncton: Dos dinoflagel·lats (en forma de fus Gyrodinium, esfèric Protoperidinium) i un tintínnid
Microzooplancton, els principals depredadors del plàncton: Dos dinoflagel·lats (en forma de fus Gyrodinium, esfèric Protoperidinium) i un tintínnid








El plàncton i la cadena alimentària
[modifica]
El plàncton és la primera baula de les cadenes alimentàries marines. El fitoplàncton és menjat pel zooplàncton i per una multitud d'organismes marins, que seran la presa de petits depredadors i aquests seran caçats pels grans depredadors. Alguns animals grans com les balenes i els taurons pelegrins s'alimenten directament de zooplàncton.
En aigües dolces, i fins i tot en aigües salobres, el fitoplàncton és una important base de les cadenes alimentàries.
En aigües particularment tèrboles carregades amb sorra o de partícules en suspensió apareixen tipus particulars de plàncton, que colonitzen les partícules en suspensió, el que permet una alta biomassa malgrat el fet que la terbolesa no permet la penetració de la llum. Aquestes aigües són generalment subjectes a l'agitació i a les fortes corrents que l'oxigenen. Un cas particular és la zona de màxima de terbolesa dels estuaris, que es mou amb les marees i l'afluència d'aigua dolça, i que serveix com a protecció i zona d'alimentació per a alevins d'algunes espècies.
La neu marina, formada pels cadàvers o els excrements del zooplàncton i que descendeixen de forma passiva al fons del mar, és de gran importància per al subministrament d'espècies d'aigües profundes i pels cicles biogeoquímics.
Algunes espècies de plàncton poden produir toxines potents (com la botulínica), que es poden incorporar en la cadena alimentària a través de mol·luscs, organismes filtradors i per alguns peixos. Aquests mateixos organismes també poden acumular toxines modificades bioacumulades en el plàncton, com el metilmercuri, quantitat que tendeix a augmentar de manera constant en els peixos depredadors i en els cetacis, i que pot ser preocupant per la salut dels consumidors humans i pels ecosistemes marins.
En determinades condicions (com una elevada aportació de nutrients, generalment materials orgànics, nitrats o fosfats), un «excés» de plàncton pot donar lloc a una situació d'eutrofització, fins i tot de distrofització, és a dir, la mort puntual o permanent de la majoria dels organismes aquàtics. L'ONU ha identificat centenars de zones mortes (Dead zone), incloses en la mar Bàltica. En aquestes àrees, l'eutrofització es pot combinar amb altres tipus de contaminació o pertorbació.
Diverses formes de plàncton (especialment de zooplàncton) tenen interès com a font de noves proteïnes per a l'alimentació humana i per a pinsos. Els problemes estan en la seva escassa digestibilitat i en el fet que un consum excessiu produiria desequilibris ecològics.[20]
El plàncton i la necromassa
[modifica]
El plàncton és l'origen d'una biomassa considerable, però també d'una necromassa que forma una part important de certs sediments (el guix és la necromassa fòssil del plàncton marí).
La sedimentació de la necromassa planctònica no és tan sols un dels embornals de carboni a nivell mundial, sinó també una de les maneres que es va desintoxicar els oceans primitius massa rics en certes sals, incloent el calci, i que va permetre la vida complexa que coneixem.
El plàncton i el clima
[modifica]El plàncton interactua amb el clima local i global. Està implicat en el cicle de carboni a través de la fotosíntesi i també, mitjançant l'emissió de molècules de sofre després de la seva mort, contribueixen a la nucleació de gotes d'aigua, és a dir a la formació de núvols i pluja. El sulfur de dimetil és dels compostos de sofre orgànics més abundants emesos a l'atmosfera i es produeix principalment en els oceans. Es degrada en l'ambient marí (principalment en diòxid de sofre, dimetilsulfòxid (DMSO), àcid sulfònic i àcid sulfúric) formant aerosols i les seves molècules actuen com a nuclis de condensació.
De mitjana, la biomassa planctònica per litre d'aigua és molt més alta en aigua freda, fins i tot sota la capa de gel, que en les aigües tropicals càlides, si estan lluny de fonts d'oligoelements com les aportacions volcàniques. Els fenòmens de surgència de les profunditats («upwelling») són la causa de la distribució massiva d'espècies de plàncton que afecten a les xarxes tròfics superiors. El canvi climàtic, que afecta els corrents oceànics i a la temperatura de l'aigua (i per tant el seu contingut d'oxigen), podria canviar la distribució i la naturalesa de les masses de plàncton, i per tant dels recursos pesquers. S'han observat canvis significatius durant gairebé un segle, però la proporció dels impactes de la pesca excessiva i la contaminació (nitrats, pesticides, metalls pesants, terbolesa, contaminació tèrmica..) en aquest fenòmens és encara difícil de determinar.
El plàncton i l'oxigen
[modifica]
El fitoplàncton, mitjançant la fotosíntesi, allibera la major part de l'oxigen del nostre planeta (el 50%), necessari també per la vida terrestre.[21]
Gràcies al fitoplàncton, l'aigua de l'oceà emmagatzema el CO₂ dissolt a la capa superficial. Es reconeix que un terç del CO₂ produït en l'atmosfera és absorbit pels mars i oceans gràcies al fitoplàncton,[22] que és tant com totes les plantes terrestres i aquàtiques, sent aquest terç el que seria el responsable de l'augment dels gasos amb efecte d'hivernacle a l'atmosfera.[23]
Més de 150 científics de 26 països han llançat una crida internacional per a aturar l'acidificació dels oceans a causa de les grans quantitats d'absorció de CO₂, ja que posa en perill els ecosistemes marins, incloent la dissolució dels esquelets calcaris de molts organismes planctònics.[Nota 1]
Història del plàncton
[modifica]Els microfòssils s'utilitzen per a estudiar com el plàncton va evolucionar dins de la biodiversitat marina. Confirmen la importància de les relacions entre el clima i el plàncton, i han demostrat que durant les grans extincions massives el plàncton també va ser fortament afectat.
En particular, un estudi recent ha comparat el contingut de microfòssils de nanoplàncton de 823 nuclis de sediments marins que provenen de 17 perforacions oceàniques fetes en els hemisferis nord i sud. Es va trobar que en el límit K-Pg (la darrera gran extinció massiva), el 93% de les espècies de nanoplàncton amb una testa calcària van desaparèixer ràpidament, amb una extinció més ràpida i més massiva en l'hemisferi nord. Aquest és un indici més per a la hipòtesi de la caiguda d'un gran asteroide en Yucatán, sobretot perquè les dates es correlacionen amb una extinció massiva d'espècies de plantes terrestres en Amèrica del Nord. Com a resultat d'aquest esdeveniment catastròfic, la diversitat del nanoplàncton en l'hemisferi nord va romandre molt més baix durant uns 40.000 anys, i va trigar gairebé 270.000 anys per a recuperar el seu nivell original. La seva diversitat és encara més important avui en dia en l'hemisferi sud. Durant aquesta extinció, el nanoplàncton fotosintètic també va quedar molt afectat, el que suggereix que l'impacte i els incendis van alliberar grans quantitats de metalls tòxics en l'aire i l'oceà,[Nota 2] i que hauria afectat més a l'hemisferi nord que a la meitat sud del planeta.[24]
Les amenaces sobre el plàncton
[modifica]
La quantitat de fitoplàncton present en superfície dels mars és seguida a gran escala per satèl·lit des de 1979. Està sotmesa a fluctuacions cícliques, en escala de dècades, relacionades a priori amb el forçant radiatiu. La durada de l'observació és encara insuficient per a predir les tendències a llarg termini, però el modelatge i l'estudi del paleoclima pot ajudar a comprendre millor les relacions entre el clima i el plàncton.
El 2006, Michael Behrenfeld (Oregon State University) mostra en la revista Nature (7 de desembre de 2006),[23] com la imatgeria per satèl·lit permet avaluar la quantitat de clorofil·la en l'aigua, i que al voltant del 60% dels mars tenien un nivell d'activitat planctònica molt baixa entre 1998 i 1999 a causa del fenomen de El Niño, abans de recuperar-se amb La Niña, i després va caure de manera constant: entre 1999 i 2005 (durant 6 anys). L'activitat planctònica sembla disminuir de forma regular i l'oceà perd de mitjana anual una capacitat d'absorció de 190 milions de tones (Mt) de carboni en comparació amb l'any anterior. Si aquesta tendència es confirma en els propers anys, el canvi climàtic podria accelerar-se. Scott Doney,[25] també en la revista Nature, va dir que, al mateix temps, la productivitat va augmentar en les latituds altes causa de l'escalfament de les aigües superficials, però que no pot compensar el dèficit de la zona tropical, el guany de la productivitat és limitat i per a un volum molt menor d'aigua.
També cal afegir els efectes de l'acidificació de l'oceà, aquells impactes pocs coneguts de la sobreexplotació, els que provoquen el blanqueig i mort dels corals, i els de l'eutrofització i la terbolesa anormal dels estuaris i de vastes àrees marines. El plàncton marí es podria veure afectat abans de 2050, o abans de 2030 en l'oceà Antàrtic. Al Mar del Nord, des de l'any 1961, la proporció de plàncton d'aigua calenta està creixent en relació a la del plàncton d'aigua freda. A més, l'acidificació afecta els esquelets calcàries del plàncton, com els d'alguns protistes, mol·luscs i crustacis.
L'escalfament global és una de les possibles amenaces, la més citada, més important que l'acidificació o la contaminació. Actua de diverses maneres:
- les «capes» d'aigua que es produeixen impliquen un menor augment de les sals nutritives per al plàncton.
- la part mòbil del plàncton contribueix indirectament a la formació de núvols i a la barreja de capes tèrmiques de diferents densitats.[2][3] Aquest cicle es pot aturar a causa que els peixos, que també contribueixen a la barreja de les capes de la superfície, són menys nombrosos i, per tant, van augmentant les zones marines mortes.
Sembla que ja han ocorregut regressions significatives de fitoplàncton, sobretot fa uns 55 milions d'anys, un període caracteritzat precisament per les taxes més altes de gasos amb efecte d'hivernacle en l'atmosfera (de causa desconeguda).
Mètodes de recollida de plàncton
[modifica]
Tradicionalment, el plàncton s'extreu amb xarxes denominades xarxes de plàncton. Aquestes xarxes estan fetes d'un cercle gran de metall al que s'afegeix una tela de niló o de seda de forma cònica que acaba amb un recipient anomenat col·lector. Hi ha diversos models de teles amb diferents mides de malla. La mida de la malla es selecciona en funció de la grandària dels organismes que es vol recollir, que és sempre menor que la mida dels organismes que es volen capturar. La xarxa es llença a l'aigua lligada a un cable. La longitud del cable desplegat permet conèixer a quina profunditat màxima és enviada la xarxa. Quan la xarxa es tira pel cable, l'aigua passa a través de les malles de la xarxa i deixar anar l'aigua; tots els organismes que tenen una grandària menor que les malles de la xarxa s'acumulen al col·lector. Normalment, l'entrada de les xarxes tenen una petita hèlix coneguda amb el nom de fluxòmetre que permet determinar la quantitat d'aigua que penetra en la xarxa. Una vegada recollida la xarxa, es desmunta el col·lector i es recull el contingut en un recipient per a ser estudiat.[26]

La xarxa de plàncton es pot utilitzar de diverses maneres. Si el vaixell està parat, la recollida es fa verticalment i les mostres proporcionen informació sobre la distribució vertical de les espècies en la columna d'aigua. Si el vaixell està en moviment, la recollida es farà horitzontalment a una profunditat determinada i proporciona informació sobre la distribució de les espècies en aquesta profunditat; en aquest cas, la xarxa té un sistema de bloqueig que s'utilitza per a evitar la contaminació de les mostres en el moment de la pujada de la xarxa. Aquests mètodes de recol·lecció de plàncton poden filtrar grans quantitats d'aigua i permet estudis en diferents tipus d'aigua, fins i tot en les zones pobres en plàncton. No obstant això, són inexactes per a l'estudi quantitatiu de plàncton. Fins i tot amb l'ús d'un fluxòmetre, segueix sent difícil estimar amb precisió la quantitat d'aigua filtrada per la xarxa i com la xarxa és arrossegada a través dels cables, no és fàcil de prendre mostres a una profunditat exacta i constant. Aquest mètode de recollida s'utilitza generalment per a l'estudi qualitatiu d'espècies, és a dir, per obtenir informació sobre la presència o absència d'una espècie en particular.
Pels estudis quantitatius precisos s'utilitzen dispositius com les ampolles de Nansen o les ampolles de Niskin. Aquests cilindres permeten recollir un volum conegut d'aigua a una profunditat precisa. L'ampolla és un cilindre de plàstic que en cada extrem té dues vàlvules que serveixen de tap. El cilindre està fixat en un cable i les vàlvules es mantenen obertes, ja sigui per un sistema de ressort o per una corda elàstica, segons el model. Es fa baixar a l'aigua amb les dues vàlvules obertes. La longitud de cable desplegat serveix per a estimar la profunditat aconseguida per l'ampolla. Quan arriba a la profunditat que es vol recollir la mostra, l'ampolla es tanca amb ajut d'un petit pes, anomenat missatger, que llisca al llarg del cable i que allibera les dues vàlvules de l'ampolla; l'aigua i el plàncton contingut en l'ampolla es queden atrapats a l'ampolla segellada i es puja a la superfície per a ser buidada. Les ampolles més modernes utilitzen electrovàlvules que poden ser accionades a una profunditat predeterminada a través d'un sensor de pressió o usant un senyal elèctric enviat per l'usuari de la superfície.[27] L'ampolla es pot usar sola o juntament amb altres ampolles per a recollir mostres simultàniament a diverses profunditats. Hi ha suports metàl·lics (la «roseta») que permeten fixar 12, 24 o 36 ampolles de mostreig[28] i que poden provocar el tancament de cada ampolla a diferents profunditats. Això fa que sigui possible recollir mostres de diferents capes de la columna d'aigua i obtenir una distribució vertical precisa de plàncton.
El plàncton i l'art
[modifica]Les formes de vegades molt geomètriques i de tipus complex de nombrosos tipus d'organismes planctònics han fascinat als científics des del seu descobriment, entre el segle xviii i el segle xix. La seva bellesa i diversitat van ser popularitzats entre el públic en general en 1904 pel best-seller del biòleg i dibuixant naturalista Ernst Haeckel titulat «Formes artístiques de la natura » (Kunstformen der Natur).
-
 Il·lustració 1 «Phaeodaria»
Il·lustració 1 «Phaeodaria» -
 Il·lustració 2 «Thalamphora»
Il·lustració 2 «Thalamphora» -
 Il·lustració 31 «Cyrtoidea»
Il·lustració 31 «Cyrtoidea» -
 Il·lustració 41 «Acanthophracta»
Il·lustració 41 «Acanthophracta» -
 Il·lustració 61 «Phaeodaria»
Il·lustració 61 «Phaeodaria» -
 Il·lustració 71 «Stephoidea»
Il·lustració 71 «Stephoidea» -
 Il·lustració 84 «Diatomea»
Il·lustració 84 «Diatomea» -
 Il·lustració 91 «Spumellaria»
Il·lustració 91 «Spumellaria»









Aquest llibre va provocar un impacte molt important en el curs de l'Art nouveau de principis del segle xx, i en particular sobre artistes com Constant Roux o René Binet, autor de la monumental porta de l'Exposició Universal de París de 1900.[29]
Notes
[modifica]- ↑ El CO₂ es combina amb una molècula d'aigua (H₂O) per a donar un ió d'hidrogen H+ i un carbonat d'hidrogen (HCO₃) que acaba descomponent-se en un carbonat (CO₃) i un altre H+. El resultat és que la concentració d'ions hidrogen en l'aigua augmenta i es fa més corrosiva, i alenteix i destrueix la síntesi de carbonat de calci, la principal matèria per a la formació de molts esquelets externs del plàncton.
- ↑ El coure és tòxic per al plàncton a molt baixes dosis (parts per bilió), però sense dubte també es van alliberar grans quantitats de níquel, cadmi i ferro i potser també crom, alumini, i especialment mercuri i plom, que tenen efectes tòxics coneguts universalment.
Referències
[modifica]- ↑ Díaz i Santos, 1998, p. 152 i 153.
- ↑ 2,0 2,1 Estudis realitzats per Katija i Dabiri al California Institute of Technology de Pasadena, recollits en la revista Nature (Nature, 29/07/2009, Nature doi:10.1038/news.2009.745)
- ↑ 3,0 3,1 «Dabiri Lab. Stanford University». Arxivat de l'original el 2011-09-02. [Consulta: 26 març 2016].
- ↑ «Online Etymology Dictionary».
- ↑ Thurman, 1997.
- ↑ Davis, 1955, p. 562.
- ↑ Karleskint, 2013.
- ↑ «plankter - marine biology». Encyclopedia Britannica.
- ↑ Mollo, 2009, p. 21-23.
- ↑ Wang, 2012, p. 71-88.
- ↑ Omori, 1992.
- ↑ Dusenbery, 2009.
- ↑ Martin, J. H.; Fitzwater, S. E. «Iron-deficiency limits phytoplankton growth in the Northeast Pacific Subarctic». Nature, 331, 6154, 1988, pàg. 341–343. Bibcode: 1988Natur.331..341M. DOI: 10.1038/331341a0.
- ↑ Boyd, P.W.; Watson, AJ; Law, CS; Abraham, ER; Trull, T; Murdoch, R; Bakker, DC; Bowie, AR; Buesseler, KO «A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by fertilization». Nature, 407, 6805 http://tass.ru/en/non-political/745635,+2000,+pàg. 695–702. DOI: 10.1038/35037500. PMID: 11048709.
- ↑ «TASS: Non-political - Scientists find traces of sea plankton on ISS surface». TASS.[Enllaç no actiu]
- ↑ Falkowski, Paul G. «The role of phytoplankton photosynthesis in global biogeochemical cycles». Photosyntheis Research, 39, 3, 1994, pàg. 235–258. DOI: 10.1007/BF00014586.[Enllaç no actiu]
- ↑ Chisholm, S.W.; Falkowski, PG; Cullen, JJ «Dis-crediting ocean fertilization». Science, 294, 5541, 2001, pàg. 309–310. DOI: 10.1126/science.1065349. PMID: 11598285.
- ↑ Aumont, O.; Bopp, L. «Globalizing results from ocean in situ iron fertilization studies». Global Biogeochemical Cycles, 20, 2, 2006, pàg. GB2017. Arxivat de l'original el 2012-11-01. Bibcode: 2006GBioC..20.2017A. DOI: 10.1029/2005GB002591 [Consulta: 26 març 2016]. Arxivat 2012-11-01 a Wayback Machine.
- ↑ Steinacher, M., et al. (2010), Steinacher, M., i al. (2010), Projected 21st century decrease in marine productivity: a multi-model analysis, Biogeosciences, 7, 979-1005
- ↑ Enciclopèdia.cat - Plàncton
- ↑ «Institut de Ciències del Mar - Plàncton». Arxivat de l'original el 2016-04-09. [Consulta: 26 març 2016].
- ↑ Comissió Oceanogràfica Intergovernamental de la UNESCO (IOC-UNESCO).
- ↑ 23,0 23,1 Michael Behrenfeld, Universitat d'Oregon, en la revista Nature (7 de desembre de 2006).
- ↑ Des traces de la météorite tueuse de dinosaures dans le nanoplancton (Les petjades del meteorit assassí de dinosaures en el nanoplàncton). Síntesi proposada per LeGéologe (francès)
- ↑ Woods Hole Oceanographic Institution (WHOI)
- ↑ Estudi del plàncton marí. Obtenció i conservació.
- ↑ «Intitut de Ciències del Mar. Operacions a coberta». Arxivat de l'original el 2016-04-09. [Consulta: 26 març 2016].
- ↑ Model 1016 Intelligent Rosette Water Sampler
- ↑ Jean-Michel Othoniel, «L'Art nouveau ou l'esthétique des courbes». France Culture.com Les jeudis de l'expo, per Elizabeth Couturier.
Bibliografia
[modifica]- Davis, C. C. The marine and fresh-water plankton (en anglès). Michigan State Univ. Press, 1955. ISBN 978-0870130168.
- Dolan, J.R; Agatha, S; Coats, D.W; Montagnes, D.J.S; Stocker, D.K. Biology and Ecology of Tintinnid Ciliates: Models for Marine Plankton (en anglès). Oxford, UK: Wiley-Blackwell, 2013. ISBN 978-0-470-67151-1.
- Dusenbery, David B. Living at Micro Scale: The Unexpected Physics of Being Small (en anglès). Cambridge, Mass: Harvard University Press, 2009. ISBN 978-0-674-03116-6.
- Haury, L. R; McGowan, J. A; Wiebe, P. H. Patterns and processes in the time-space scales of plankton distribution. Pattern in plankton communities (en anglès). Nova York: (eds J. H. Steele), Plenum Press, 1978, p. 227-327.
- Karleskint, George; Turner, Richard; Small, James. Introduction to Marine Biology. Cap 17, The Open Sea (en anglès). Brooks/Cole, 2013. ISBN 978-1-133-36446-7.
- Kiørboe, Thomas. A Mechanistic Approach to Plankton Ecology (en anglès). Princeton, N.J.: Princeton University Press, 2008. ISBN 978-0-691-13422-2.
- Kirby, Richard R. Ocean Drifters: A Secret World Beneath the Waves (en anglès). Regne Unit: Studio Cactus Ltd, 2010. ISBN 978-1-904239-10-9.
- Mollo, Pierre; Thomas-Bourgneuf, Maëlle. L'enjeu plancton, l'écologie de l'invisible (en francès). Ed. Charles Léopold Mayer, 2009. ISBN 978-2843771477.
- Mollo, Pierre; Noury, Anne. Le Manuel du plancton (en francès). Ed. Charles Léopold Mayer, 2013.
- Omori, M; Ikeda, T. Methods in Marine Zooplankton Ecology (en anglès). Malabar, USA: Krieger Publishing Company, 1992. ISBN 0-89464-653-2.
- Ott, Jörg. Meereskunde. Einführung in die Geographie und Biologie der Ozeane. 2 Auflage (en alemany). Stuttgart: UTB, 1996. ISBN 3-8252-1450-8.
- Sardet, Christian. Plankton: Wonders of the Drifting World (en anglès). The University of Chicago Press, 2015. ISBN 978-0-226-26534-6.
- Thurman, H. V. Introductory Oceanography (en anglès). New Jersey, USA: Prentice Hall College, 1997. ISBN 0-13-262072-3.
- Wang, G; Wang, X; Liu, X; Li, Q. Diversity and biogeochemical function of planktonic fungi in the ocean (en anglès). Raghukumar, 2012.
Vegeu també
[modifica]- Acidificació de l'oceà
- Anabaena
- Bomba biològica
- Flor d'aigua
- Plèuston
- Zona morta
- Zooplàncton gelatinós
Bibliografia
[modifica]- Díaz, J.; Santos, T. Zoología: Aproximación evolutiva a la diversidad y organización de los animales (en castellà). Editorial Síntesis, 1998. ISBN 84-7738-591-2.
Enllaços externs
[modifica]- Vida al mar - Bloc amb informació sobre el plàncton a la Mediterrània. (català)