Phycobilisomes are light-harvesting antennae that transmit the energy of harvested photons to photosystem II and photosystem I in cyanobacteria and in the chloroplasts of red algae and glaucophytes.[1][2][3] They were lost during the evolution of the chloroplasts of green algae and plants.[3]
| Phycobilisome protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 The layout of protein subunits in a phycobilisome. | |||||||||
| Identifiers | |||||||||
| Symbol | Phycobilisome | ||||||||
| Pfam | PF00502 | ||||||||
| InterPro | IPR012128 | ||||||||
| SCOP2 | 1cpc / SCOPe / SUPFAM | ||||||||
| |||||||||
General structure
editPhycobilisomes are protein complexes (up to 600 polypeptides) anchored to thylakoid membranes. They are made of stacks of chromophorylated proteins, the phycobiliproteins, and their associated linker polypeptides. Each phycobilisome consists of a core made of allophycocyanin, from which several outwardly oriented rods made of stacked disks of phycocyanin and (if present) phycoerythrin(s) or phycoerythrocyanin. The spectral property of phycobiliproteins are mainly dictated by their prosthetic groups, which are linear tetrapyrroles known as phycobilins including phycocyanobilin, phycoerythrobilin, phycourobilin and phycobiliviolin. The spectral properties of a given phycobilin are influenced by its protein environment.[4]
Function
editEach phycobiliprotein has a specific absorption and fluorescence emission maximum in the visible range of light. Therefore, their presence and the particular arrangement within the phycobilisomes allow absorption and unidirectional transfer of light energy to chlorophyll a of the photosystem II. In this way, the cells take advantage of the available wavelengths of light (in the 500–650 nm range), which are inaccessible to chlorophyll, and utilize their energy for photosynthesis. This is particularly advantageous deeper in the water column, where light with longer wavelengths is less transmitted and therefore less available directly to chlorophyll.
The geometrical arrangement of a phycobilisome is very elegant in an antenna-like assembly. It results in 95% efficiency of energy transfer.[5]
Evolution and diversity
editThere are many variations to the general phycobilisome structure. Their shape can be hemidiscoidal (in cyanobacteria) or hemiellipsoidal (in red algae). Species lacking phycoerythrin have at least two disks of phycocyanin per rod, which is sufficient for maximum photosynthesis.[6]
The phycobiliproteins themselves show little sequence evolution due to their highly constrained function (absorption and transfer of specific wavelengths).[citation needed] In some species of cyanobacteria, when both phycocyanin and phycoerythrin is present, the phycobilisome can undergo significant restructuring as response to light color. In green light the distal portions of the rods are made of red colored phycoerythrin, which absorbs green light better. In red light, this is replaced by blue colored phycocyanin, which absorbs red light better. This reversible process is known as complementary chromatic adaptation. It is the component of photosynthetic system of cyanobacteria, as a particle with which various structures are linked (i.e. thylakoid membrane, etc.).[citation needed]
Applications
editPhycobilisomes can be used in prompt fluorescence Archived 2018-03-18 at the Wayback Machine,[7][8] flow cytometry,[9] Western blotting and protein microarrays. Some phycobilisomes have an absorption and emission profile similar to Cy5, allowing them to be used in many of the same applications. They can also be up to 200 times brighter and with a larger Stokes shift, providing a larger signal per binding event. This property allows the detection of low-level target molecules[9] or rare events.
-
 Excitation and emission spectra of a phycobilisome from a blue-green alga.
Excitation and emission spectra of a phycobilisome from a blue-green alga. -
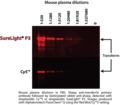 Phycobilisome versus cyanine dye detection capabilities in Western blot application.
Phycobilisome versus cyanine dye detection capabilities in Western blot application.


References
edit- ^ Kawakami K, Hamaguchi T, Hirose Y, Kosumi D, Miyata M, Kamiya N, Yonekura K (2022). "Core and rod structures of a thermophilic cyanobacterial light-harvesting phycobilisome". Nature Communications. 13. doi:10.1038/s41467-022-30962-9. PMC 9205905. Art. No. 3389.
- ^ Chang L, Liu X, Li Y, Liu CC, Yang F, Zhao J, Sui SF (2015). "Structural organization of an intact phycobilisome and its association with photosystem II". Cell Research. 25 (6): 726–737. doi:10.1038/cr.2015.59. PMC 4456626. PMID 25998682.
- ^ a b Stadnichuk IN, Kusnetsov VV (2021). "Endosymbiotic Origin of Chloroplasts in Plant Cells' Evolution". Russian Journal of Plant Physiology. 68 (1): 1–16. doi:10.1134/S1021443721010179. S2CID 255012748.
- ^ Singh NK, Sonani RR, Rastogi RP, Madamwar D (2015). "The phycobilisomes: an early requisite for efficient photosynthesis in cyanobacteria". EXCLI Journal. 14: 268–89. doi:10.17179/excli2014-723. PMC 4553884. PMID 26417362.
- ^ Glazer AN (June 1985). "Light harvesting by phycobilisomes". Annual Review of Biophysics and Biophysical Chemistry. 14: 47–77. doi:10.1146/annurev.bb.14.060185.000403. PMID 3924069.
- ^ Lea-Smith DJ, Bombelli P, Dennis JS, Scott SA, Smith AG, Howe CJ (June 2014). "Phycobilisome-Deficient Strains of Synechocystis sp. PCC 6803 Have Reduced Size and Require Carbon-Limiting Conditions to Exhibit Enhanced Productivity". Plant Physiology. 165 (2): 705–714. doi:10.1104/pp.114.237206. PMC 4044857. PMID 24760817.
- ^ Zoha SJ, Ramnarain S, Morseman JP, Moss MW, Allnutt FT, Rogers YH, Harvey B (1999). "PBXL Fluorescent Dyes for Ultrasensitive Direct Detection". Journal of Fluorescence. 9 (3): 197–208. doi:10.1023/A:1022503600141. S2CID 12373519.
- ^ "MicroPlate Detection comparison between SureLight®P-3L, other fluorophores and enzymatic detection" (PDF). Technical Bulletin 3. Columbia Biosciences. 2010. Archived from the original (PDF) on 18 March 2018.
- ^ a b Telford WG, Moss MW, Morseman JP, Allnutt FC (August 2001). "Cyanobacterial stabilized phycobilisomes as fluorochromes for extracellular antigen detection by flow cytometry" (PDF). Journal of Immunological Methods. 254 (1–2): 13–30. doi:10.1016/s0022-1759(01)00367-2. PMID 11406150. Archived from the original (PDF) on 2018-03-18. Retrieved 2014-06-07.
Further reading
edit- Zheng L, Zheng Z, Li X, Wang G, Zhang K, Wei P, Zhao J, Gao N (2021). "Structural insight into the mechanism of energy transfer in cyanobacterial phycobilisomes". Nature Communications. 12. doi:10.1038/s41467-021-25813-y. PMC 8448738. Art. No. 5497.
External links
edit- Phycobilisomes at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Columbia Biosciences - Phycobilisome Resource