
Homo
Pour les articles homonymes, voir Homo (homonymie).

Homo est le genre qui réunit Homo sapiens et les espèces apparentées. Il apparait à la fin du Pliocène ou au début du Pléistocène, selon l'attribution des plus anciens fossiles faite par les paléoanthropologues. Depuis quelque 2,5 millions d'années, le genre Homo a produit un buissonnement d'espèces, en raison de sa dispersion géographique précoce dans tout l'Ancien Monde et de son développement dans des niches écologiques variées.
Toutes les espèces du genre Homo sont aujourd'hui éteintes, sauf Homo sapiens. Les dernières espèces apparentées, Homo floresiensis, Homo luzonensis, Homo denisovensis et Homo neanderthalensis, ont disparu il y a entre 50 000 et 30 000 ans.
Étymologie
[modifier | modifier le code]Homo, en latin, signifie « homme », « être humain », au sens générique, tandis que vir désigne l'« homme » au sens de « mâle », opposé à femina, « femme ». Le mot dérive du latin archaïque hĕmo, lui-même issu de l'indo-européen *dʰǵʰm̥mō (« [chose / fils] de la terre »).
Carl von Linné crée en 1758 le genre Homo, dans lequel il place deux espèces, Homo sapiens (encore nommé Homo diurnus) et Homo troglodytes (aussi appelé Homo nocturnus ou Homo sylvestris), ce dernier décrit par des voyageurs résultant d'un amalgame entre l'orang-outan asiatique et le chimpanzé africain[1].
Classification phylogénétique
[modifier | modifier le code]La classification phylogénétique moderne positionne le genre Homo dans une succession de groupes en cascade, depuis la famille des hominidés (Hominidae) jusqu'à la sous-tribu des Hominina, qui inclut aussi, selon le consensus actuel, les genres éteints Sahelanthrope, Orrorin, Ardipithèque, Kenyanthrope, Australopithèque et Paranthrope.
Phylogénie des genres actuels d'hominidés, d'après Shoshani et al. (1996)[2] et Springer et al. (2012)[3] :
| Hominidae |
| ||||||||||||||||||
L'espèce de grand singe génétiquement la plus proche d'Homo sapiens est le Chimpanzé. Les deux espèces partagent 98,8 % de leurs gènes[4]. Homo sapiens a notamment deux chromosomes de moins que le chimpanzé[a], mais les deux génomes conservent beaucoup de similitudes[b].
Selon la plupart des paléoanthropologues, la divergence entre Hominina (lignée des humains) et Panina (lignée des chimpanzés) remonterait à au moins 7 millions d'années[5]. Les généticiens ont longtemps avancé le chiffre de 5 à 6 millions d'années, mais sur la base d'une horloge moléculaire dont le calibrage était fondé sur des taux de mutation qui ont été révisés à la baisse en 2016[6].
Émergence
[modifier | modifier le code]Le fossile le plus ancien découvert à ce jour et attribué au genre Homo (mis au jour en 2013 en Éthiopie et désigné sous le code LD 350-1) remonte à la fin du Pliocène, il y a environ 2,8 millions d'années[7]. Il est cependant trop fragmentaire pour avoir pu recevoir une dénomination.
Homo habilis et Homo rudolfensis sont les deux espèces décrites les plus anciennes du genre Homo, remontant à environ 2,4 millions d'années en Afrique. L'espèce les précédant dans l'évolution n'est pas encore identifiée. Quoique de nombreux chercheurs pensent que le genre Homo est probablement issu d'une espèce du genre Australopithèque, il demeure possible que ces deux genres soient des développements parallèles issus d'un ancêtre commun.
L'apparition du genre Homo est souvent interprétée comme le résultat d'une divergence phylogénétique sous la pression de l'environnement. À partir de 3 millions d'années, la tendance globale au refroidissement et à l'assèchement du climat, avec réduction du couvert forestier et accroissement des habitats ouverts, auraient poussé des Hominina à exploiter de nouvelles niches écologiques, comprenant un complément de ressources alimentaires à base de viande et de graisse. Obtenue par le charognage ou la chasse de petits animaux, cette alimentation plus riche aurait notamment favorisé un début d'accroissement du volume cérébral[8]. Le paléoanthropologue français Yves Coppens a constaté cet assèchement du climat dans la stratigraphie de la vallée de l'Omo, au sud de l'Éthiopie, sur la période allant de 3 à 2 millions d'années (transition Plio-Pléistocène). Il en a tiré la théorie dite de l'Évènement de l'(H)Omo, qui succède depuis les années 2000 à la précédente version de sa théorie, dite de l'East Side Story[9].
Une étude publiée en 2020 semble confirmer ce lien entre l'origine de l'Homo et les changements climatiques et environnementaux entre 3 et 2,5 Ma. Elle présente de nouveaux résultats qui mettent en lumière l'interaction entre la tectonique, la migration des bassins, les changements fauniques d'une part et le sort de l'Australopithecus afarensis et l'évolution de l'Homo de l'autre. Les travaux sur le terrain au nouveau site de Mille-Logya dans l'Afar, en Éthiopie, datés entre 2,914 et 2,443 Ma, fournissent des preuves géologiques de la migration nord-est du bassin de Hadar, étendant la documentation de ce bassin lacustre jusqu'à Mille-Logya. Les chercheurs ont identifié trois nouvelles unités fossilifères, suggérant un changement faunique in situ dans cet intervalle. Alors que la faune de l'unité plus ancienne est comparable à celle de Hadar et Dikika, les unités plus jeunes contiennent des espèces qui indiquent des conditions plus ouvertes ainsi que des restes d'Homo. Cela suggère que celui-ci a émergé des Australopithèques pendant cet intervalle ou s'est dispersé dans la région dans le cadre d'une faune adaptée à des habitats plus ouverts[10].
Goulot d'étranglement
[modifier | modifier le code]Une étude publiée en 2023 utilise un nouveau modèle coalescent pour prédire la taille passée de la population humaine à partir de plus de 3 000 génomes humains actuels. Le modèle détecte une réduction de la taille de la population des ancêtres Homo d'environ 100 000 à environ 1 000 individus. Ce déclin semble avoir coïncidé avec un changement climatique majeur, notamment des périodes prolongées de refroidissement du climat, et des événements de spéciation ultérieurs. Les ancêtres humains auraient donc connu un grave goulot d'étranglement démographique avec environ 1 280 individus reproducteurs il y a environ 930 000 à 813 000 ans. Ce goulot d’étranglement aurait duré environ 117 000 ans et aurait rapproché les ancêtres humains de l’extinction. Il est conforme à un écart chronologique important dans les archives fossiles africaines et eurasiennes disponibles[11]. D’autres études avaient déjà constaté une baisse importante du nombre d’humains. Néanmoins, cette enquête génétique « exclut tous les groupes humains qui pouvaient vivre à cette époque mais qui ne sont pas nos ancêtres directs »[12].
Cette situation démographique aurait divisé les humains en petits groupes distincts qui, progressivement, ont pu développer des différences anatomiques suffisamment importantes pour donner naissance à des populations survivantes distinctes. C’est ainsi que seraient apparus les Néandertaliens, les Dénisoviens et l'Homo sapiens[11].
Des chercheurs émettent cependant des doutes vis-à-vis des dates (celles-ci étant déterminées par le choix arbitraire d'une durée de génération de 24 ans) et de l'estimation de la taille de population efficace (en) proposées par l'étude. Pour Thierry Grange, généticien spécialiste des populations anciennes, la différence de dates « peut-être 200 000 ans en plus ou en moins ». Chris Stringer, paléoanthropologue au Natural History Museum (Londres) estime peu probable une si faible population sur près de 100 000 ans[12]. La paléogénéticienne Céline Bon rappelle que cette étude est basée sur la théorie de la coalescence qui repose sur des hypothèses simplificatrices discutables (âge de reproduction, tous les couples avec le même nombre d’enfants…)[13].
Caractères anatomiques propres au genre Homo
[modifier | modifier le code]Plusieurs caractères morphologiques (qui s'expriment variablement selon les espèces) permettent de définir le genre Homo.
Bipédie
[modifier | modifier le code]Caractères communs avec les Hominina
[modifier | modifier le code]Les Hominina se caractérisent notamment par leur bipédie. Celle-ci inclut les éléments suivants :
- quatre courbures sur la colonne vertébrale[14] ;
- trou occipital avancé à la base du crâne, ce qui est en relation avec une position verticale ;
- bassin court et évasé qui permet une meilleure stabilité debout[14] ;
- fémurs obliques par rapport à la verticale, ce qui permet aux pieds d'être à l'aplomb du centre de gravité.
Caractères propres du genre Homo
[modifier | modifier le code]
Les représentants du genre Homo présentent en plus les caractères suivants :
- membres postérieurs sensiblement plus longs que les membres antérieurs ;
- hallux parallèle aux autres orteils ;
- existence d'une voûte plantaire.
Il convient de noter qu'Homo habilis présente une morphologie intermédiaire qui ne répond pas complètement aux caractéristiques listées dans ce deuxième paragraphe. Le squelette post-crânien d'Homo rudolfensis demeure quant à lui inconnu.
Un nombre croissant de chercheurs ajoutent aux 3 caractéristiques précédentes l'aptitude à la course, à partir de l'analyse chez les spécimens fossiles des os du pied et d'un certain nombre d'adaptations morphologiques lisibles sur les ossements fossiles. Cette position s'appuie largement sur la théorie du coureur de fond.
Évolution crânienne
[modifier | modifier le code]- capacité crânienne de plus de 550 cm3, supérieure aux Australopithèques et en augmentation progressive au cours du Pléistocène ;
- aplatissement de la face par la réduction du prognathisme (mâchoire avancée) ;
- réduction des muscles de l'appareil masticatoire ;
- réduction de la denture, avec une décroissance de la taille des molaires de M1 à M3 (elles sont de taille croissante chez les Australopithèques)[5].
Capacité vocale
[modifier | modifier le code]- descente progressive du larynx, libérant ainsi la capacité de moduler des sons articulés[5] ;
- l'os hyoïde, qui joue un rôle dans l'expression vocale, est présent chez l'Homme de Néandertal ; sa présence ou non chez les autres espèces humaines non sapiens est inconnue.
Thorax
[modifier | modifier le code]- cage thoracique en forme de tonneau (celle des Australopithèques est plus conique)[14] ;
- deux côtes flottantes, non rattachées au sternum (contre une seule chez les Australopithèques)[14].
Autres caractères anatomiques
[modifier | modifier le code]- pouce de la main plus long que chez les Australopithèques, permettant une meilleure saisie de précision.
Caractères culturels propres au genre Homo
[modifier | modifier le code]Quelques activités ou capacités culturelles paraissent spécifiques au genre Homo :
- utilisation d'outils lithiques avec l'apparition de galets taillés (Oldowayen), puis l'amélioration de l'outillage (Acheuléen) ;
- consommation régulière de viande, acquise par la chasse ou par le charognage[15] ;
- capacité à vivre à des latitudes un peu plus septentrionales (en Europe et en Asie) que les autres genres d'hominidés ;
- utilisation occasionnelle du feu dès 1,5 Ma, sans capacité de le produire à volonté avant environ 400 000 ans[16].
Espèces connues du genre Homo
[modifier | modifier le code]La taxonomie des Hominina préhumains et des espèces du genre Homo est en évolution permanente. Choisir de faire de l'un d'eux une nouvelle espèce en lui attribuant un nom binominal, choisir de modifier leur définition selon l'évolution de leur hypodigme, sont des pratiques régulières qui apparaissent au fil des découvertes et des débats. Pour illustrer ces doutes, on préfère parfois accoler les périodes d'existence connues des différents Hominina et espèces humaines sur une frise chronologique, sans lien de parenté :

Les spécialistes du Musée de l'Homme, à Paris, reconnaissent aujourd'hui 14 espèces distinctes au sein du genre Homo[17], auxquelles on peut ajouter l'Homme de Denisova, qui a été identifié en 2010 par son génome mais n'a pas encore été décrit sur la base de fossiles.
Tableau comparatif
[modifier | modifier le code]La variabilité des critères dimensionnels peut être extrêmement élevée au sein d'une même espèce, à la fois en fonction de facteurs géographiques et du facteur temps[18]. De nombreuses espèces d'hominidés ne sont pas suffisamment échantillonnées pour tirer des conclusions sur les schémas de dimorphisme sexuel dans leurs populations fossiles ; dans ce cas ce dimorphisme sexuel est déduit par inférence à partir de celui de populations actuelles d'hommes et de singes : la taille et le poids des sujets estimés féminins sont toujours moindres, mais dans des proportions variables selon les espèces[19].
- Les espèces notées en gras indiquent l'existence d'un nombre important de fossiles découverts.
- CE : Coefficient d'encéphalisation
| Espèces | Période (Ma) | Localisation | Fossiles | Date de découverte |
Publication du nom |
|---|---|---|---|---|---|
| Homo sp. (espèce encore non dénommée) |
2,8[7] | Éthiopie (Afrique) | 1 mandibule | 2013 | 2015 |
| H. habilis | 2,3 - 1,4 | Afrique orientale | peu de postcrânien | 1960 | 1964 |
| H. rudolfensis | 2,4 - 1,8 | Afrique orientale | pas de postcrânien | 1972 | 1986 |
| H. gautengensis | 2,0 - 0,8 | Afrique australe | fragmentaires | 1952 | 2010 |
| H. georgicus | 1,77 | Géorgie | 5 crânes | 1991 | 2002 |
| H. ergaster | 1,9 - 1,0 | Afrique, Moyen-Orient | nombreux | 1971 | 1975 |
| H. erectus | 1,0 - 0,1 | Asie (Java, Chine) | nombreux | 1891 | 1894 |
| H. antecessor | 0,85 | Espagne | 1 site (Gran Dolina) | 1994 | 1997 |
| H. heidelbergensis | 0,7 - 0,3 | Europe | peu de postcrânien | 1907 | 1908 |
| H. rhodesiensis | 0,6 - 0,2 | Afrique | peu de postcrânien | 1921 | 1921 |
| H. naledi | 0,3[20] | Afrique australe | au moins 15 individus | 2013 | 2015 |
| H. floresiensis | 0,19 - 0,05 | Indonésie | 1 individu semi-complet | 2003 | 2004 |
| H. luzonensis | 0,07 - 0,05 | Philippines | fragmentaires | 2007 | 2019 |
| H. neanderthalensis | 0,45 - 0,03 | Europe, Moyen-Orient, Asie Centrale, Altaï |
nombreux | 1856 | 1864 |
| H. denisovensis | 0,45 - 0,04 | Sibérie, Asie orientale | très peu | 2008 | 2010 |
| H. sapiens | 0,3[21] - présent | Afrique, puis Monde | actuel | 1758 |
Liste
[modifier | modifier le code]



Arago 21 (Homme de Tautavel)

- Homo antecessor n'a été trouvé que sur un site de la Sierra d'Atapuerca, en Espagne. C'est la plus ancienne espèce identifiée en Europe, les fossiles plus anciens étant trop fragmentaires pour pouvoir être attribués.
- Homo denisovensis est un groupe apparenté à l'Homme de Néandertal. Identifié pour la première fois dans l'Altaï, en Sibérie, grâce à une analyse génétique, il aurait vécu plus largement dans toute l'Asie orientale, où de nombreux fossiles du Paléolithique moyen déjà connus sont en attente d'attribution. Il se serait éteint vers la même époque que Néandertal.
- Homo erectus a été trouvé à Java et en Chine. Il est peut-être issu d'Homo ergaster ou d'une espèce humaine antérieure ayant évolué en Asie. Les derniers Homo erectus connus sont datés d'environ 110 000 ans, à Java.
- Homo ergaster, apparu en Afrique de l'Est, est la première espèce humaine dotée d'un squelette post-crânien proche de l'Homme moderne, montrant une bipédie permanente et une aptitude à la course. Le crâne conserve en revanche une morphologie archaïque.
- Homo floresiensis est une espèce de petite taille qui serait endémique de l’île de Florès, en Indonésie, où elle se serait développée à l'écart des espèces humaines voisines, l'ile de Florès n'ayant jamais été connectée au continent par voie terrestre lors des cycles glaciaires du Pléistocène.
- Homo gautengensis est une espèce archaïque, décrite en 2010 sur la base de fossiles fragmentaires trouvés depuis 1952 en Afrique du Sud et qui avaient été attribués auparavant à Homo habilis ou à Homo ergaster.
- Homo georgicus, découvert en 1991 en Géorgie, est à ce jour le plus ancien représentant du genre Homo attesté hors d'Afrique. L'attribution de certains fossiles plus anciens trouvés en Chine reste débattue par les chercheurs. Homo georgicus présente une morphologie intermédiaire entre Homo habilis et Homo ergaster.
- Homo habilis est l'une des plus anciennes espèces connues du genre Homo. Sa capacité crânienne montre une première augmentation, encore modeste, par rapport aux Australopithèques. Homo habilis aurait eu une bipédie affirmée mais avec une capacité arboricole encore présente.
-
 Homo habilis
Homo habilis
OH 24 -
 Homo rudolfensis
Homo rudolfensis
KNM-ER 1470 -
 Homo floresiensis
Homo floresiensis
Liang Bua 1 (LB 1) -
 Homo ergaster
Homo ergaster
KNM-ER 3733 -
 Homo rhodesiensis
Homo rhodesiensis
Homme de Kabwe -
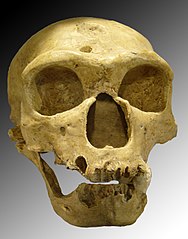 Homo neanderthalensis, La Chapelle-aux-Saints 1
Homo neanderthalensis, La Chapelle-aux-Saints 1




.jpg)

- Homo heidelbergensis est une espèce qui rassemble la plupart de ceux des fossiles européens du Pléistocène moyen qui ne sont pas attribués à l'Homme de Néandertal. Il est considéré comme l'ancêtre probable des Néandertaliens et des Dénisoviens.
- Homo luzonensis, à l'image de l'Homme de Florès, serait une espèce endémique de l'ile de Luçon, aux Philippines, où elle se serait développée à l'écart des espèces humaines voisines, l’île de Luçon n'ayant jamais été connectée au continent par voie terrestre lors des cycles glaciaires du Pléistocène.
- Homo naledi est une espèce proche des Australopithèques par son modeste volume endocrânien, mais que sa gracilité et sa complète bipédie rapprochent des autres espèces humaines connues[22].
- Homo neanderthalensis a un crâne un peu plus volumineux qu'Homo sapiens, mais allongé vers l'arrière, large et peu élevé. La face présente des bourrelets sus-orbitaires marqués. L'Homme de Néandertal est particulièrement robuste et trapu, avec des membres courts, ce qui est une constitution adaptée au froid. Les derniers Hommes de Néandertal connus ont été découverts au sud de la péninsule Ibérique et en Crimée.
- Homo rhodesiensis est une espèce morphologiquement proche d'Homo heidelbergensis, mais qui s'est développée en Afrique. Elle montre un volume endocrânien assez élevé tout en ayant conservé un fort torus sus-orbitaire. Son statut d'ancêtre potentiel d'Homo sapiens est âprement discuté.
- Homo rudolfensis serait plus grand et plus lourd qu'Homo habilis. Sa denture est plus puissante, ce qui laisse supposer une alimentation orientée plutôt vers des végétaux coriaces.
- Homo sapiens possède un crâne globulaire, avec un front élevé, une face réduite, et une mince arcade sourcilière. La symphyse mandibulaire présente un menton. Son squelette est plus gracile que celui des espèces antérieures. Originaire d'Afrique, il se diffuse sur toute la planète où il remplace les espèces humaines locales. Il est l'auteur de l'art préhistorique du Paléolithique supérieur.
Phylogénie
[modifier | modifier le code]
Phylogénie des espèces récentes du genre Homo, d'après Strait, Grine & Fleagle (2015)[23], et Meyer & al. (2016)[24] :
| Homo |
| |||||||||||||||||||||||||||||||||
À droite, modélisation de la distribution géographique de quelques espèces du genre Homo durant les deux derniers millions d'années.
L'axe horizontal représente la localisation géographique, tandis que l'axe vertical représente le temps en millions d'années. La surface bleue indique la présence de certaines espèces sur un continent et sur une période donnée.
Les hommes modernes se sont propagés depuis l'Afrique vers toutes les régions du globe[25].
Périmètre
[modifier | modifier le code]Certains chercheurs, à la suite du paléoanthropologue anglais Bernard A. Wood, professeur d'anthropologie à l'Université George-Washington (États-Unis), estiment qu'Homo habilis et Homo rudolfensis devraient être exclus du périmètre du genre Homo, lequel commencerait alors il y a environ 2 millions d'années avec l'apparition d'Homo ergaster. Ils constatent en effet en Afrique à partir de cette époque les traces d'une amplification importante des activités de boucherie et la disparition de plusieurs espèces de prédateurs concurrentes du genre Homo, concomitamment avec la naissance de l'industrie acheuléenne, dont la première manifestation est datée de 1,76 million d'années au Kenya[26]. De plus, les fossiles d'Homo habilis et d'Homo rudolfensis ont révélé une bipédie encore incomplète, suggérant que ces deux espèces étaient encore partiellement arboricoles. Elles n'auraient pas été aptes à la course bipède, caractère jugé par certains comme nécessaire à l'appartenance au genre Homo.
Migrations hors d'Afrique
[modifier | modifier le code]Le consensus scientifique veut que Homo soit apparu en Afrique.
Migrations d'Afrique vers l'Eurasie
[modifier | modifier le code]Homo georgicus est attesté en Géorgie il y a 1,77 Ma, associé à une industrie lithique de type oldowayen. La plus ancienne trace d'occupation humaine en Europe a été trouvée à Kozarnika, en Bulgarie. Elle est datée de 1,6 Ma[réf. souhaitée]. Les gisements de la région d'Orce, en Espagne, ont livré des vestiges fossiles et lithiques datés d'environ 1,4 Ma.
Homo ergaster, considéré comme l'auteur de l'industrie acheuléenne en Afrique à partir de 1,76 Ma (sites de Kokiselei 4, au Kenya, et de Konso Gardula 6, en Éthiopie), serait tôt sorti de son berceau africain en diffusant l'Acheuléen au Moyen-Orient (site d'Ubeidiya, en Israël, daté de 1,4 Ma) et en Inde (site d'Attirampakkam, dans le Tamil Nadu, daté de 1,5 Ma). L'Homme de Denizli, un fossile trouvé en Turquie et daté entre 1,2 et 1,6 Ma, est attribué à Homo erectus par la paléoanthropologue Amélie Vialet[27],[28] (qui relève aussi des similitudes avec Homo ergaster)[29].
Le début tardif de l’Acheuléen en Europe occidentale et méridionale, il y a environ 800 000 ans, pourrait être associé à la sortie d'Afrique d'une population devenue en Europe Homo heidelbergensis, probable ancêtre de l'Homme de Néandertal[30]. L'Homme de Denisova, apparenté à l'Homme de Néandertal, pourrait s'être étendu de l'Ouest vers l'Est, occupant au Paléolithique moyen la Sibérie et l'Asie orientale.
Les différentes sorties d'Afrique de groupes humains archaïques auraient pu avoir lieu à des périodes favorables sur le plan climatique, en particulier lors des périodes interglaciaires, qui faisaient à la fois remonter la zone tropicale vers le Nord et augmenter les précipitations, réduisant ainsi les barrières géographiques que constituent les zones désertiques[c].
Thèse des premiers Homo apparus en Asie
[modifier | modifier le code]La découverte de 2009 à 2017 de vestiges archéologiques dans la chaine des Siwaliks, au pied de l'Himalaya, en Inde, par Anne Dambricourt-Malassé et Claire Gaillard, pourrait remettre en cause l'origine africaine du genre Homo, ou reculer sensiblement la date de sa première sortie d'Afrique. L'équipe de chercheurs franco-indienne a trouvé à Masol, dans le nord-ouest de l'Inde, des vestiges datés de 2,6 millions d'années habituellement attribués au genre Homo. Quatre os d'animaux présentent des marques de découpe effectuées par un tranchant lithique[31],[32],[33]. En 2017 a été trouvé sur le même site le premier outil taillé encore en place dans la stratigraphie, également daté de 2,6 Ma. Ces découvertes rendent envisageable l'existence d'un foyer d'hominisation en Asie du Sud, à moins qu'il convienne de reporter la première sortie d'Afrique du genre Homo au-delà de 2,6 Ma, c'est-à-dire environ 800 000 ans plus tôt que la date avancée jusqu'à présent.
D'autres vestiges lithiques ou fossiles, découverts en Israël, au Pakistan et en Chine, contribuent à montrer une sortie d'Afrique précoce, antérieure à 2,1 millions d'années[34],[35].
Homo sapiens
[modifier | modifier le code]Des vestiges fossiles d'Homo sapiens ont été découverts en Israël dès les années 1930, dans ce qui est considéré comme des sépultures, dans la grotte d'Es Skhul, datés d'environ 118 000 ans, et dans la grotte de Qafzeh, datés d'environ 92 000 ans[36]. Plus récemment, un demi-maxillaire d'Homo sapiens, daté d'environ 185 000 ans, découvert en 2002 dans la grotte de Misliya (Israël) et publié en 2018, a fait remonter la première sortie d'Afrique d'Homo sapiens 70 000 ans plus tôt que le laissaient penser les précédentes découvertes[37],[38]. Une mandibule fragmentaire attribuée à Homo sapiens a été découverte en 2008 en Chine, dans la grotte de Zhiren, située dans la province du Guangxi. Elle est datée d'environ 110 000 ans[39].
Selon les études génétiques réalisées sur les populations humaines actuelles, la dernière sortie d'Afrique par une population d'Homo sapiens aurait eu lieu il y a environ 55 000 ans. Deux hypothèses ont été avancées pour le passage de l'Afrique vers l'Eurasie : par l'isthme de Suez puis le corridor levantin[40], ou en traversant la mer Rouge par le détroit de Bab-el-Mandeb jusqu'à la péninsule Arabique[41].
Hybridation
[modifier | modifier le code]Les différentes espèces humaines qui ont souvent coexisté n'étaient pas isolées sur le plan reproductif. Les équipes de l'Institut Max-Planck d'anthropologie évolutionniste, à Leipzig (Allemagne), ont notamment mis en évidence depuis 2010 plusieurs flux d'hybridation anciens entre les Homo sapiens, les Néandertaliens et les Dénisoviens, ainsi qu'avec d'autres espèces encore non identifiées en Afrique et en Asie, dans des proportions qui restent toutefois marginales (de 0 à 4 % de la partie variable du génome de l'Homme moderne[d]). Les populations actuelles non subsahariennes possèdent notamment en moyenne 1,8 % de gènes néandertaliens, répartis à différents endroits de leur génome[42],[43].
Quelques ossements fossiles témoignent d'une hybridation récente. Analysé en 2015, l'ADN d'un Homo sapiens découvert en Roumanie et datant de 37 800 à 42 000 ans avant le présent comporte 7,3 % de gènes néandertaliens, et la longueur des segments de gènes néandertaliens indique que son ancêtre Néandertal remontait à seulement quatre à six générations[44]. En 2018, l'ADN d'un fragment osseux trouvé dans la grotte de Denisova, celui d'une adolescente morte vers 13 ans il y a environ 90 000 ans[e], montre qu'elle était l'hybride d'une mère néandertalienne et d'un père dénisovien[45]. C'est la première preuve directe d'un croisement entre espèces humaines.
Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]- Les deux paires de chromosomes {2p, 2q} de l'ancêtre commun d'Homo sapiens et du Chimpanzé ont fusionné en la paire de chromosomes {2} chez l'Homme, qui a par suite 46 chromosomes, alors que le Chimpanzé a conservé les deux paires de chromosomes {2p, 2q} de l'ancêtre commun et a donc toujours 48 chromosomes.
- En plus des chromosomes sexuels {X, Y} qui sont très voisins, il subsiste treize autres paires de chromosomes {3, 6, 7, 8, 10, 11, 12, 14, 16, 19, 20, 21, 22} qui semblent pratiquement identiques entre les deux espèces, ainsi que six paires qui sont restées proches à la suite d'insertions {1}, d'inversions {4, 5, 17} et de délétions {13, 18} relativement simples et facilement identifiables, mais deux paires de chromosomes {9, 15} ont été plus profondément différenciées à la suite de mutations complexes, probablement échelonnées dans le temps au sein des sous-tribus Hominina et/ou Panina.
- Voir par exemple l'intervalle humide que représente le subpluvial néolithique (Sahara vert).
- La partie variable du génome de l'espèce humaine, c'est-à-dire les gènes qui peuvent varier d'un individu à l'autre, représente environ 1 pour mille du génome global.
- Il s'agit du fragment osseux dénommé Denisova-11. L'adolescente a été surnommée Denny.
Références
[modifier | modifier le code]- Claude-Louis Gallien, Homo. Histoire plurielle d'un genre très singulier, Presses universitaires de France, , p. 87.
- (en) J. Shoshani, C. P. Groves, E. L. Simons et G. F. Gunnell, « Primate phylogeny : morphological vs. molecular results », Molecular Phylogenetics and Evolution, vol. 5, no 1, , p. 102-54 (PMID 8673281, lire en ligne).
- (en) Mark S. Springer, Robert W. Meredith et al., « Macroevolutionary Dynamics and Historical Biogeography of Primate Diversification Inferred from a Species Supermatrix », PLoS ONE, vol. 7, no 11, , e49521 (ISSN 1932-6203, PMID 23166696, PMCID 3500307, DOI 10.1371/journal.pone.0049521, lire en ligne).
- (en) David R. Begun, The real Planet of the Apes : A new Story of human Origins, 2015
- Pascal Picq, Premiers hommes, éd. Flammarion, Paris, 2016
- (en) Priya Moorjani, Carlos Eduardo G. Amorim, Peter F. Arndt et Molly Przeworski, « Variation in the molecular clock of primates », PNAS, vol. 113, no 38, , p. 10607–10612 (lire en ligne)
- Pascaline Minet, « Paléontologie : une mandibule africaine vieillit le genre humain de 400 000 ans », Le Monde, 4 mars 2015.
- (en) Peter B. deMenocal, « African climate change and faunal evolution during the Pliocene-Pleistocene », Earth and Planetary Science Letters, vol. 220, nos 1-2, , p. 3-24 (DOI 10.1016/S0012-821X(04)00003-2).
- Yves Coppens, L'évènement de l'(H)Omo, Dossiers du CNRS, lire en ligne.
- (en) Zeresenay Alemseged et al., Fossils from Mille-Logya, Afar, Ethiopia, elucidate the link between Pliocene environmental changes and Homo origins, Nature Communications, volume 11, Article numéro: 2480 (2020
- (en) Wangjie Hu, Ziqian Hao, Pengyuan Du et al., Genomic inference of a severe human bottleneck during the Early to Middle Pleistocene transition, Science, Vol. 381, Numéro 6661, pp. 979-984, 31 août 2023, DOI: 10.1126/science.abq7487
- Sébastian Seibt, L'humanité a-t-elle failli disparaître il y a 900 000 ans ?, france24.com, 1er septembre 2023
- Céline Bon, « L'espèce humaine a-t-elle frôlé l'extinction il y a 900 000 ans ? », sur museedelhomme.fr,
- (en) Carol Ward, « Early Hominin Body Form », sur CARTA - The Upright Ape : Bipedalism and Human Origins, .
- Jean-Jacques Hublin, Quand d'autres hommes peuplaient la Terre : combien d'espèces ?, conférence au Musée de l'Homme, 19 octobre 2018, voir la vidéo en ligne.
- (en) Lyn Wadley, « The Origin and Development of Fire Technology in Africa », sur CARTA - The Perspective from Africa, .
- Dominique Grivaud-Hervé, « Combien d'espèces Homo sont à ce jour recensées par les chercheurs ? », sur museedelhomme.fr, (consulté le ).
- Manuel Will, Adrián Pablos et Jay T. Stock, « Long-term patterns of body mass and stature evolution within the hominin lineage », Royal Society Open Science, vol. 4, no 11, , p. 171339 (DOI 10.1098/rsos.171339, lire en ligne, consulté le )
- (en) Brian Villmoare, Kevin G. Hatala et William Jungers, « Sexual dimorphism in Homo erectus inferred from 1.5 Ma footprints near Ileret, Kenya », Scientific Reports, vol. 9, no 1, , p. 7687 (ISSN 2045-2322, DOI 10.1038/s41598-019-44060-2, lire en ligne, consulté le )
- (en) Paul H.G.M. Dirks, Lee R. Berger et al., « The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa », eLife, vol. 6, no e24231, (DOI 10.7554/eLife.24231, lire en ligne)
- Hervé Morin, « La découverte qui bouleverse l’histoire d’« Homo sapiens » », Le Monde.fr, (ISSN 1950-6244, lire en ligne, consulté le )
- Hervé Morin, « "Homo Naledi", une découverte qui laisse perplexe », Le Monde.fr, (ISSN 1950-6244, lire en ligne, consulté le ).
- [Strait, Grine & Fleagle 2015] (en) David Strait, Frederick Grine et John Fleagle, « Analyzing Hominin Phylogeny : Cladistic Approach », dans Winfried Henke & Ian Tattersall, Handbook of Paleoanthropology, (ISBN 9783642399787, lire en ligne), p. 1989-2014.
- [Meyer et al. 2016] (en) Matthias Meyer et al., « Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins », Nature, vol. 531, no 7595, , p. 504-507 (DOI 10.1038/nature17405, résumé).
- (en) C. Stringer, « What makes a modern human », Nature, vol. 485, no 7396, , p. 33–35 (DOI 10.1038/485033a).
- Jean-Jacques Hublin, chaire de Paléoanthropologie au Collège de France, L'émergence du genre Homo, 14 octobre 2014 (voir la vidéo).
- Amélie Vialet, Gaspard Guipert, Mehmet Cihat Alçiçek et Marie-Antoinette de Lumley, « La calotte crânienne de l’Homo erectus de Kocabaş (Bassin de Denizli, Turquie) », L'Anthropologie, le site de Kocabaş, bassin de Denizli, Anatolie, Turquie, vol. 118, no 1, , p. 74–107 (ISSN 0003-5521, DOI 10.1016/j.anthro.2014.01.003, lire en ligne, consulté le )
- « La Turquie et l’expansion des premiers Hommes. », sur Muséum national d'Histoire naturelle (consulté le ).
- Amélie Vialet, Sandrine Prat, Une contribution de la Turquie au carrefour eurasiatique, Colloque au Collège de France, 3-4 juillet 2018, voir la vidéo en ligne.
- Jean-Jacques Hublin et Bernard Seytre, Quand d'autres hommes peuplaient la terre. Nouveaux regards sur nos origines, Flammarion, , p. 87.
- « Des hominidés en Inde il y a 2,6 million d'années », sur hominides.com (consulté le ).
- (en) Anne Dambricourt Malassé, Mukesh Singh, Baldev Karir et Claire Gaillard, « Anthropic activities in the fossiliferous Quranwala Zone, 2.6 Ma, Siwaliks of Northwest India, historical context of the discovery and scientific investigations », Comptes Rendus Palevol, origines de l’homme dans le sous-continent Indien, vol. 15, nos 3–4, , p. 295–316 (DOI 10.1016/j.crpv.2015.06.004, lire en ligne, consulté le ).
- (en) Claire Gaillard, Mukesh Singh, Anne Dambricourt Malassé et Vipnesh Bhardwaj, « The lithic industries on the fossiliferous outcrops of the Late Pliocene Masol Formation, Siwalik Frontal Range, northwestern India (Punjab) », Comptes Rendus Palevol, origines de l’homme dans le sous-continent Indien, vol. 15, nos 3–4, , p. 341–357 (DOI 10.1016/j.crpv.2015.09.017, lire en ligne, consulté le ).
- (en) Hao Li, ChaoRong Li et Kathleen Kuman, « Longgudong, an early Pleistocene site in Jianshi, South China, with stratigraphic association of human teeth and lithics », Science China Earth Sciences, vol. 60, no 3, , p. 452-462 (lire en ligne).
- (en) Colin Barras, « Tools from China are oldest hint of human lineage outside Africa », Nature, (ISSN 0028-0836 et 1476-4687, DOI 10.1038/d41586-018-05696-8, lire en ligne, consulté le ).
- Bernard Vandermeersch, « Qafzeh, histoire des découvertes », Bulletin du Centre de recherche français à Jésuralem, vol. 18, , p. 8-19 (lire en ligne).
- François Savatier, « Homo sapiens est arrivé au Levant bien avant les néandertaliens », Pour la science, no 485, , p. 6-7.
- (en) Israel Hershkovitz, Gerhard W. Weber, Rolf Quam, Mathieu Duval, Rainer Grün et al., « The earliest modern humans outside Africa », Science, vol. 359, no 6374, , p. 456-459 (DOI 10.1126/science.aap8369).
- (en) Yanjun Cai, « The age of human remains and associated fauna from Zhiren Cave in Guangxi, southern China », Quaternary International, (DOI 10.1016/j.quaint.2015.12.088).
- (en) Robin Derricourt, « Getting “Out of Africa” : Sea Crossings, Land Crossings and Culture in the Hominin Migrations », Journal of World Prehistory, vol. 19, no 2, , p. 119–132 (DOI 10.1007/s10963-006-9002-z).
- (en) S. Mithen, M. Reed, « Stepping out: a computer simulation of hominid dispersal from Africa », J Hum Evol., vol. 43, no 4, , p. 433-462.
- Science, Comparative Genomics : the Neandertal Genome
- (en) « Welcome to the family, Homo sapiens neanderthalensis », sur New Scientist, (consulté le ).
- (en) Qiaomei Fu, Mateja Hajdinjak, Oana Teodora Moldovan, Silviu Constantin, Swapan Mallick et al., « An early modern human from Romania with a recent Neanderthal ancestor », Nature, vol. 524, , p. 216-219 (DOI 10.1038/nature14558)
- (en) Matthew Warren, « Mum’s a Neanderthal, Dad’s a Denisovan : First discovery of an ancient-human hybrid - Genetic analysis uncovers a direct descendant of two different groups of early humans », Nature, vol. 560, , p. 417-418 (DOI 10.1038/d41586-018-06004-0).
Voir aussi
[modifier | modifier le code]Bibliographie
[modifier | modifier le code]- Dominique Grimaud-Hervé, Frédéric Serre, Jean-Jacques Bahain et al., Histoire d'ancêtres : La grande aventure de la Préhistoire, Paris IVe, Errance, coll. « Guides de la préhistoire mondiale », , 5e éd., 144 p. (ISBN 978-2-87772-590-3)
Articles connexes
[modifier | modifier le code]- Histoire évolutive de la lignée humaine
- Australopithèque
- Paranthrope
- Hominisation
- Bipédie
- Évolution du cerveau
Liens externes
[modifier | modifier le code]- (en) Hominid species
- (en) Lancelet and Ancestral Body (article : évolution des chordés, patrimoine génétique, découvertes récentes), sur penichefossil.net [PDF]